
北の海のかけら/後編

★表紙は、pixabayからダウンロードしたカムチャッカ半島の景色
★前編はこちら
*****
ミトコンドリアハプログループ(mtハプロ)とY染色体ハプログループ(Yハプロ)を調べてもらった話の続き。
その後、追加レポートの作成(私たち現生人類であるホモサピエンスと交配していたネアンデルタール人のゲノムを自分はどれぐらい持ってるかとか)をお願いしたり、書籍や論文を読んでいろいろ調べたりしていたのと、どうやらこれはミソジニーの一種のように思えるのですが、Yハプロに異常に執着する考え方が世の中にはあるらしく(書籍でも批判されてた)、正直なところとても気持ち悪いので、記事の続きはどう書いたらいいものかと悩んでました。
しかし、その人の特性にmtハプロは影響する可能性はある(エネルギー産生器官であるミトコンドリアをそのまま引き継いでいるわけだし)だろうけれど、Y染色体って受精後の生物学的な男性化のトリガーに一瞬なるだけで別に何の役にも立ってないし(だからどんどん短くなってるし、なんならX染色体に組み換えて乗せてしまえそう)、なんなのかなあれは。
それはともかく結果発表です。
*****
mtハプロ:G1a1a
mtハプロはG1グループでした。a1aは、バリエーション(変異)を示す枝番です。
日本人のうちG1のmtハプロを持つ人は、約3.8%とのことです。Gグループ全体(G1とG2の合計)では7%弱ぐらいらしい。日本で一番多いのはDグループで、だいたい40%ぐらいの人はDグループに属しているそう。それに比べれば少数派。
現生人類の発祥の地であるアフリカからスタートしてGに至る移動経路は、現在のmtハプログループの分布から推測してこんな感じ。
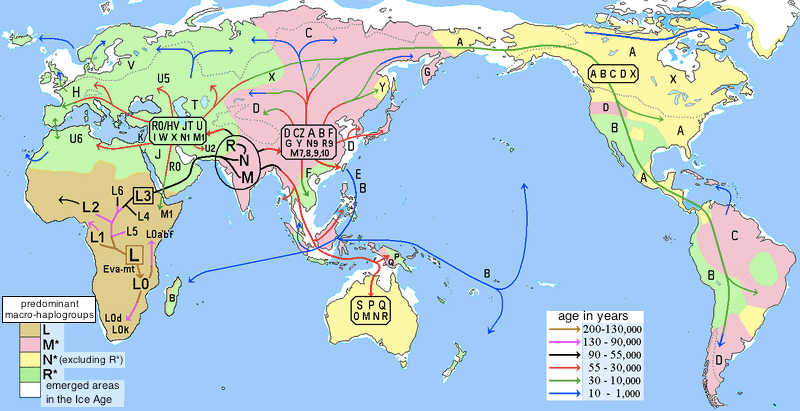
アフリカのLからスタートしてアラビア半島を経てユーラシア(南アジア)に入り、Nと分岐してMになって東進、その後、ひたすら北東に進んで北東シベリアの辺りからカムチャッカ半島に至るルートです(ユーラシアの東の端にひっそりとGが分布しているのがわかります?)。ただし、Aグループのようにベーリング海峡を通って北米大陸に渡ることはなくて、北への移動が遅かったのか、はたまた、Aグループが渡ったときには凍結して地続きだったベーリング海峡はすでに海になってしまっていたのでしょうか。
グループGのうち、その一部が後に南下してサハリン/北海道ルートか、沿海州/朝鮮半島ルートを経由して日本列島に入って定着し、その末裔が私ということになります。このルート沿いには、mtハプロがGグループの人が比較的多いようで、日本だと北海道に多いらしい。
さて、G1であることがわかってからカムチャッカ半島やその対岸である沿海州に住んでいる少数民族の人たちの顔をいろいろ眺めていたのですが、彼らの風貌は日本でも普通に見かける感じの風貌でして、なんとなく母の姉妹と似ている人もちらほらいる。
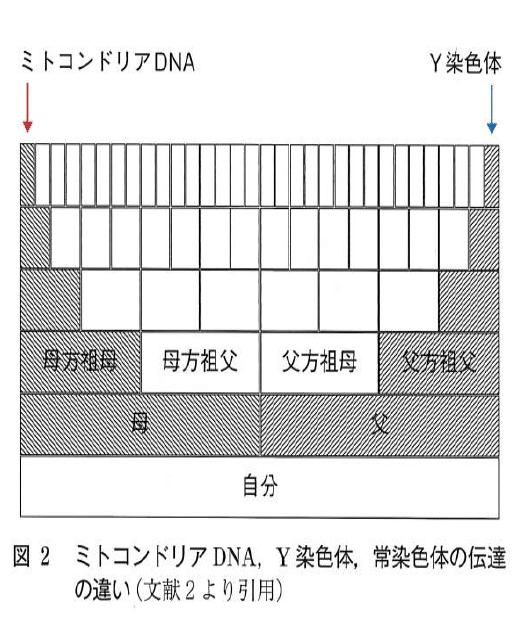
ただし、当たり前ながら容姿のブレは結構大きくて(母の姉妹の間でも全然違うわけで)、第一、前編に書いたように「ハプログループで見えるものは母mにつながる系統と、父fにつながる系統だけで、その他の大勢の母父たち(下にある図2の白い部分)のことは見えないので、調べたその人自身が母m系統と父f系統の和のみからできているわけではない」わけです。顔が似てるどうこうは気のせいですね。
とはいえ、カムチャッカ半島には鼻の形が私とよく似ている人がちらほらといて、あまり日本では見かけない形なのですが(若い頃にいろいろ言われたこともある)、もしかしてこの鼻は寒冷地仕様だったのかな?
寒冷地仕様といえば、Gグループのミトコンドリアの特徴として、エネルギー生成効率が他のグループのものよりも高い可能性があるという論文を見つけました。論文によれば、有酸素運動系競技のアスリートのmtハプロがGである確率は有意に高いんだって。Gグループが形成された極寒の地では、エネルギー生成効率が高くないと生きていけなかったのかもしれないですね。
実際にどうかと言われれば寒いのが好きで(当然、冬が一番好き)、変に暖房とか入っていると暑くてたまらなく感じる方なので、確かにエネルギー生成効率は高いのかもしれない。もっとも、西日本育ちなので、暑いのも結構平気なのは平気だけれど。
でもやっぱり冬が好き。
*****
Yハプロ:D1a2a
YハプロはD1グループでした。a2aは枝番です。実は、a2aの後に枝番がさらに6桁も続く(“a”だけではなく初めて“b”も登場する)のですが省略します。なんでそんなに細かいのかな?
日本人男性のうちD1のYハプロを持つ人は3割から4割とのことです。最も割合が大きいのはOグループで6割ほどらしい。
現生人類の発祥の地であるアフリカからスタートしてD1に至る移動経路は、現在のYハプログループの分布から推測してこんな感じ。

小さくて見えづらいですが、東北から北海道あたりにくすんだ緑色の部分がありますよね。そこがD1a2a。東アジア部分の移動経路の拡大図もWikipediaにありまして、こんな感じ。

これを見ると、出アフリカの後、北ルートであるシルクロード辺りを通過して中国の北方を通り、朝鮮半島を経由して日本列島に入ってきたようです。
D1a2aが日本列島に入って縄文人集団を形成した後、中国大陸から稲作を携えてOグループの人たちが渡来系弥生人として流入し、日本列島に拡がっていったそうで。
日本列島に到達するまでの区間にmtハプロほどのダイナミックさが感じられなくて少々残念ですが、到達後はどうしていたんでしょうか。私の遥か祖先は、Oグループの流入によって、一度、東北や北海道まで北上した後に西日本に戻ってきたのか、あるいは、西日本に留まったまま、流入してきた中国大陸からの弥生人たちと溶け込んでいったのか。どっちなんでしょうね?
*****
さて、前回、あまり何も考えずに、
なんと!
渡来系弥生人の系統ではなかったのです。
とか書いてしまいましたが、この図を思い出してください。
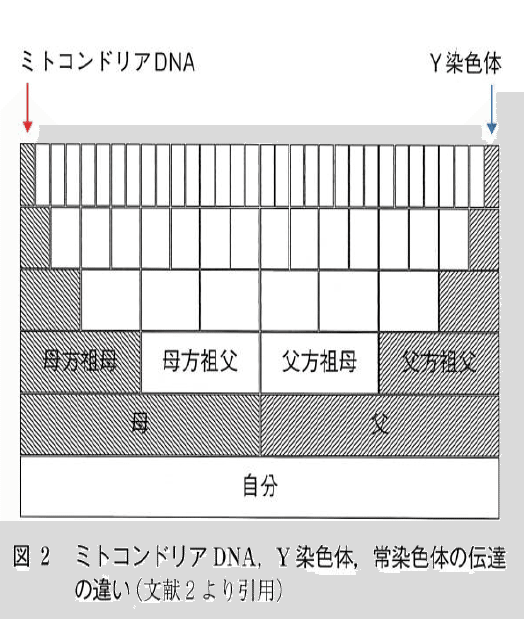
「ハプログループでは、ほとんどの母父は見えない」
mtハプロとYハプロのどちらも縄文系であるからといって、最終的に出来上がったもの(私)については正直なところ何も関係なさそうです。
そこで、縄文人(縄文人の定義が不明だけれど、遺跡などで発見された平均的な縄文人という意味なのかな?)と一致する遺伝子をどの程度持っているかをレポートしてもらいました。


あははは。平均ですね。最頻値(分布曲線のピークの横軸値)そのもの。日本でごくごく一般的な縄文/弥生の混合ですよね、これは。
ところで頻度分布を見ると、縄文人と一致する遺伝子を全く持っていない人も2%弱いるらしく、この集団は渡来系弥生人の末裔と言っていいんでしょうかね。
*****
さて、今回の追加レポートで一番楽しみにしていたネアンデルタール人に一致する遺伝子をどの程度持っているかについて。
結果発表


こちらも平均。最頻値よりはちょっと多い。単純平均(赤い破線のところ)は、この頻度分布だとあまり関係ないですね(一致する数がとても多い少数の人が単純平均値を高くしている)。
ちなみに、ネアンデルタール人に由来する遺伝子は、状況によって良い方向に働くことも、悪い方向に働くこともあるようです。
*****
どういう理屈になっているのかはよくわかりませんが、追加レポートでは私の由来する民族構成度も表示されます。
結果は、
日本民族:91%
朝鮮民族:8%
不明:1%
でした。
朝鮮民族の割合は、YハプロD1a2aの移動経路を考えると妥当であるように思えます。そして、もしかして、mtハプロG1a1aの移動経路はサハリン/北海道ルートではなく、沿海州/朝鮮半島ルートだったのかも。もっとも、上の図の白い部分の人たち(見えない無数の母父たち)によるんですけどね。
意外なのは中国民族の割合がゼロなことで、確かにmtハプロもYハプロも中国経由のハプログループではありませんが、上の図の白い部分の人たち(見えない無数の母父たち)にもいなかったのかな。それとも、いたけれど、数が少なかったので遺伝子上の痕跡が見えなくなったとかでしょうか?
そして私の遺伝子の約1%を占めている民族不明の要素
どういう経緯で痕跡を残すことになったのかはまったくわからないのですが、私の祖先にいる、この1%を与えてくれた人たちのことが大好きです。
1%のあなたたちがいてくれたからこそ、今の私がある。
ありがとう