
【自然科学】地球年表書いてみたい(ほぼ自分用メモ帳)

7月24日 21時の金曜ロードショーはジュラシック・ワールド
長いので飛ばしながら見てね。
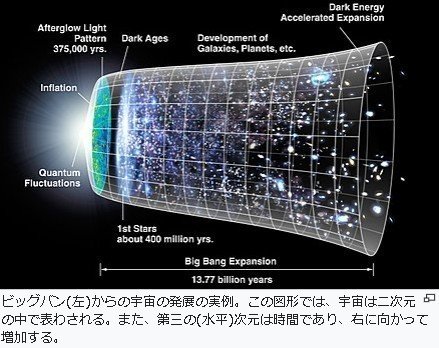
138億年前
宇宙誕生
ビッグバン
太陽は、過去の超新星爆発で散らばった星間物質がふたたび集まって形成された種族Iの星であると考えられ、太陽系は46億年前に形成され始めたとされる。また、太陽系には鉄や金、ウランといった重元素が多く存在している。これらの重元素の成因としては、質量の大きな高温の星の内部での元素合成とその後に発生する超新星爆発によって作られ、宇宙空間にまき散らされた、という過程が最も可能性が高いシナリオだからである。なお2014年現在では、金やウランなど原子番号40以上の元素の成因を超新星爆発に求める説は懐疑的となり、中性子星合体による可能性が高くなってきている
46億年前
地球誕生
先カンブリア時代(46億 - 5億4200万年前)

冥王代
45億年前ー40億年前
地球と他の星の誕生の同一性
地球と隕石から「放射性物質ヨウ素129を起源とするキセノン129」が検出される。ヨウ素129は半減期が1600万年しかない短寿命の放射性物質であり、この元素が形成される超新星爆発のあと1億年程度でほとんど消滅する。すなわち地球や隕石が形成される少し前に、近傍で超新星爆発があったとされる[6]。これは地球と隕石が同一箇所で同一時期に形成された可能性が高いことを示す。また隕石に含まれる各元素の量(元素存在度)を調べると、太陽の光球の元素存在度と良く一致する。地球は中心部に鉄主体の核を持つため、地上で手に入る地殻の元素存在度は上記太陽や隕石と異なるが、最初に地球ができたときの成分は太陽や隕石と同じであったと考えられる。このように太陽系の星は同時に同一の原料から誕生したとされる。
マグマオーシャン ジャイアントインパクト直後の原始地球の姿

微惑星の衝突が減ってくると原始地球の表面や大気が冷えていった。
大気に含まれていた大量の水蒸気は冷やされ豪雨となり地上に降り注いだ。
そして地表に海ができた。
こうして地球をおおっていた厚い雲は姿を消した。
大気の主成分は二酸化炭素となり、酸性だった海も陸地の物質が溶け込むことで中和し大気中の二酸化炭素を海に溶け込ませることが可能になった。
温室効果をもたらす二酸化炭素が海に溶け込むことによって地球の気温は安定していった。
もし雨が降らなければ、大気最上部の水蒸気が太陽からの紫外線により酸素と水素に分解され、軽い水素は宇宙に放出され、地球から水蒸気がなくなっていた怖れがあった。

太陽系の隕石や月の岩石の生成年代から、この頃、原始地球が形成されたと考えられている。地球の形成は太陽系の形成と進化での説明が詳しい。
水、アンモニア、メタンなどの水素化合物が凝集し固体となるのに充分な低温となる主たる恒星からの距離を凍結線と呼び、太陽系の場合、凍結線は約2.7AUであり、小惑星帯の辺りになる。原始太陽系星雲内で凍結線よりも温度の低いところでは、降着による微惑星および惑星の生成が、これらの固体となった粒子によって起こりやすくなる。したがって凍結線は惑星の質を地球型と木星型に分ける境界になる。H2O(水)が昇華する温度がおよそ170Kであり、凍結線の内側ではH2O(水)は水蒸気に、外側では氷になり、そのため凍結線の内側では地球のような岩石の惑星が形成され、その外側には氷の惑星ができる
また、地球の形成位置は、後に生命を育むことができる液体の水を保持することができるハビタブルゾーンに位置していた。
月の形成時期も、45億5000万年前とされる。月の形成仮説の1つとして、地球に原始惑星が衝突して形成されたとするジャイアント・インパクト説がある。
地球が誕生して間もない45億4000万年前から44億4000万年前のマントルに由来する溶岩が、カナダ・バフィン島とグリーンランド西部で見つかったと、米カーネギー研究所の研究チームが2010年8月『ネイチャー』に発表した。
地球大気の歴史については、確証は得られていないが、以下のようなことが考えられている。地球が誕生した46億年前頃の原始大気は、主にヘリウムと水素からなり、高温高圧だった。これは現在の太陽の大気と似た成分である。これらの軽い成分は、原始太陽の強力な太陽風によって数千万年のうちにほとんどが吹き飛ばされてしまったと考えられている。やがて、太陽風は太陽の成長とともに次第に弱くなってくる。この頃には、地表の温度が低下したことで地殻ができ、地殻上で多くの火山が盛んに噴火を繰り返していた。この噴火にともなって、二酸化炭素とアンモニアが大量に放出された。水蒸気と多少の窒素も含まれていたが、酸素は存在しなかった。この原始大気は二酸化炭素が大半を占め、微量成分として一酸化炭素、窒素、水蒸気などを含む、現在の金星の大気に近いものであったと考えられている。100気圧程度と濃く、高濃度の二酸化炭素が存在した。地球が十分に冷却されていなかった時期の原始大気には大量の水蒸気が含まれていたと考えられる
44億年前 - 現在、知られている最古の岩石鉱物が現れる。
西オーストラリア州のジャック・ヒルズで発見されたジルコン粒子のうち最古の物(44億400万±800万年前)
ジルコン粒子の中にダイヤモンドが含まれていることが、2007年に明らかになった。このジルコン粒子年代値は、30億年 - 42.5億年前である
41億年前 - 38億年前 後期重爆撃期には、木星型惑星の移動による重力の釣り合いの変化によって、多数の小天体が内部太陽系に侵入し、それ以前の元々小惑星帯にあった物質が枯渇して現在の状態になったこれが引き金となって、月に多くの衝突クレーターが形成され、地球・水星・金星・火星といった岩石惑星も多くの天体衝突を受けたと考えられている。天体衝突に由来する月面の溶融岩石の大部分がこの短い期間に作られたと示されている。
40億年前 - 原始海洋ができた。花崗岩(カナダ北部のアカスタ片麻岩)ができた。プレートができた
古い変成岩に含まれる堆積岩の痕跡などから、43 - 40億年前頃に海洋が誕生したとみられる。この海洋は、原始大気に含まれていた水蒸気が、火山からの過剰な噴出と温度低下によって凝結して、雨として降り注いで形成されたものであった。初期の海洋は、原始大気に含まれていた亜硫酸や塩酸を溶かしこんでいたため酸性であったが、陸地にある金属イオンが雨とともに流れ込んである程度中和されたと考えられている。ある程度中和されると二酸化炭素が溶解できるようになるため、大量の二酸化炭素を吸収していった。地球全体は還元的な雰囲気下にあり、鉄は2価鉄のイオンとして溶解していた。水蒸気が紫外線を受けて光分解することで酸素が生成されてはいたが、2価鉄が3価鉄への酸化により発生した酸素がすぐに吸収されたため、大気中にはほとんど残らなかった。
太古代
40億年前ー25億年前
原核生物から真核単細胞生物が現れるまで
30億年より前の海水温度は60-120℃という高温
太古代を通じて大気中には酸素はなく窒素と二酸化炭素が主体であった。27億年前頃から大陸周辺にシアノバクテリアが形成した大規模なストロマトライトから酸素が放出され始める。

40億年前
生命の誕生

38億年前 - 現在、知られている最古の堆積岩が現れる。
西グリーンランドのイスア地方(約38億年前)の表成岩やイサック片麻岩複合体など。堆積作用があったことから、この頃には海が存在していたと考えられている。
38億年前(±3億年) - 真正細菌(バクテリア)と古細菌(アーキア)の出現。 → 共通祖先。
共通祖先に近い原始的な生物は好熱性を示すものが多く見られる。例えば、真正細菌の根に一番近いのは超好熱性水素細菌である。古細菌でも根に近いものは好熱性のものに占められている。最も早い推計では、タンパク質アミノ酸配列の置換速度から、共通祖先は42億年前にいたとする研究がある。
35億年前 - 生物活動の化石証拠
地球上での最古の化石(西オーストラリア・ピルパラ地域からのバクテリアの化石)。メタン生成の痕跡(西オーストラリア・ピルパラ地域から、同位体比率異常のメタン)
32億年前
光合成をする生物が現れる。藍藻(シアノバクテリア)。

ストロマトライトとして痕跡を残した。ストロマトライトは藍藻(シアノバクテリア)の活動で形成された岩石。また、これよりも古い時代とする説もある。遅くとも32億年前までには光合成をする生物が現れ、海中に酸素を供給しはじめた。
二酸化炭素光合成を行う生物が誕生すると、それらは二酸化炭素を酸素に変換するようになる。さらに、二酸化炭素が生物の体内に有機物として蓄積されるようになり(炭素固定)、長い時間をかけて過剰な炭素は化石燃料、生物の殻からできる石灰岩などの堆積岩といった形で固定される。
27億年前 - シアノバクテリアが大量発生。
この頃のシアノバクテリアの化石が大量に見つかっている。酸素の供給量が増加。
原生代
25億年前から5億4,100万年前
この時大陸がはじめて安定した。そして光合成によりエネルギーと酸素を作り出す微生物、シアノバクテリアが誕生した。
古生物学の状況証拠によると、18億年前頃の一日は20時間であり、一年は400日であったと推定されている
25億年前 - 縞状鉄鉱層が形成される。
シアノバクテリアの活動で海中の酸素量が増加し、海中の2価の鉄イオンが3価鉄に酸化して沈殿したため形成される。縞状鉄鉱層の形成がおよそ19億年前まで続き、これ以後は形成されなくなる。
24億 - 22億年前 - 現在分かっている最古の氷期。→ ヒューロニアン氷期
22億2000年前 - マクガニン氷河時代、全球凍結
20数億年前? - 大気中の酸素の増加。酸素は初期の生物の大量絶滅と酸素を効果的に利用した生物のさらなる進化を導いた。
海中の鉄イオン濃度が低下し、海中の鉄イオンが酸化し尽くされると縞状鉄鉱層の形成も停止し、余剰となった海中の酸素が大気中にも多く供給されるようになった。
大気中の酸素は紫外線と反応しオゾンをつくった。酸素濃度が低かったころは地表にまで及んでいたオゾン層は、濃度の上昇とともに高度が高くなり現在と同じ成層圏まで移動した。これにより地表に到達するDNAを破壊する有害な紫外線が減少し、生物が陸上にあがる環境が整えられた。
21億年前(±6億年) - ミトコンドリア、葉緑体等に相当する生物と共生した真核生物の出現。
20億年前
- 現存する最古かつ最大の小惑星衝突 → フレデフォート・ドーム

20億 - 19億年前 - 最初の超大陸(ヌーナ大陸)出現か?
大陸移動説によれば、大陸は数億年程度の周期で離散集合を繰り返していると考えられ、この頃、ヌーナ大陸と名づけられた超大陸が出現したと考えられている。
約20億 - 10億年前 - 酸素濃度が現代の1/100以上のレベルとなり大きくは増加しなかった。
約10億 - 7億年前 - ロディニア大陸誕生
10億 - 6億年前 - この頃、多細胞生物が出現したと考えられている。多細胞生物は原口 (生物学)の獲得により強力な捕食能を有するに至った。
8億5000万年前頃 - この頃の1年は約435日。
ストロマトライトからの計測結果による。
10億年前のジャイアント・インパクト説

現在ではジャイアントインパクト説は否定されている
理由 月の成分がほぼ地球と同等である事(ジャイアントインパクトではぶつかった星の成分も混入する)
仮説 複数衝突説の登場

数値計算によると、地球に火星サイズの天体1個が衝突して月は形成されたとするシナリオでは、月の成分の5分の1は地球に由来し、残る5分の4は衝突した天体に由来することになる。しかしながら、実際には地球と月の成分構成(例えば酸素同位体比)がほぼ同一であることから、ジャイアント・インパクト仮説には物質科学的な問題点も存在している。この問題を解決するシナリオとして、イスラエル・ワイツマン科学研究所のラルカ・ルフらは複数衝突説を提唱している。
複数衝突説は、月は巨大衝突説が唱えるように1回の大規模衝突によって形成されたのではなく、複数の天体衝突の末に月が形成されたとする説である。この説では、微惑星の小さな衝突が20回程度繰り返され、衝突のたびに原始地球の周囲に残骸の輪が形成され、小衛星となり、こうした小衛星が合体することで最終的に月が形成されたとする。複数衝突説では、放出物質の地球由来物質の寄与が大きい衝突も考慮できる点や、月組成が多数の小衛星の組成を平均化した組成となることから、地球と月の物質科学的類似性の問題は緩和される。また、多様な衝突シナリオを考慮できる
8億 - 6億年前 - 大規模な氷河時代であったとされる。
7億年前 - スターチアン氷河時代、全球凍結
6億5000万年前 - マリノアン氷河時代、全球凍結
全球凍結(スノーボールアース)仮説。
7億1700万年前ー6億8000万年前
スノーボールアース
スターチアン氷期

6億4000万年前ー6億3500万年前
スノーボールアース
マリノアン氷期



氷から解けた酸素で海洋生物は生き延びたと思われる。
6億 - 5億5千万年前
先カンブリア時代
エディアカラン
多細胞生物が現れる。

エディアカラ生物群は、地球全体が氷に覆われていた時期(スノーボールアース)の直後に出現し、その大部分がカンブリア紀の始まる前に絶滅した。
エディアカラ生物群に属するいくつかの生物は、従来カンブリア紀に入ってから突然出現したと考えられていた動物群の直接の祖先であるとされるようになってきている。
古生代カンブリア紀
最初の甲殻類
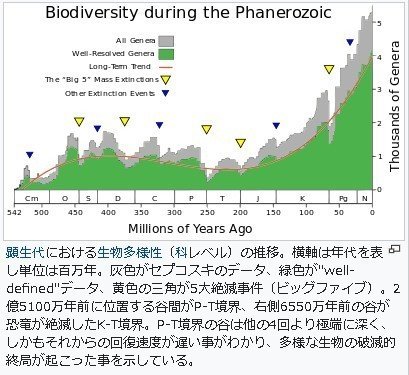
5大インパクト(5つの大量絶滅)
多細胞生物が現れたエディアカラン以降、5度の大量絶滅
O-S境界 4億4380万年前のオルドビス紀とシルル紀
F-F境界 3億7220万年前の後期デボン紀のフラニアン期とファメニアン期
P-T境界 2億5,100万年前の古生代オエルム紀とトリアス紀
T-J境界 2億130万年前の三畳紀トリアス紀とジュラ紀
K-Pg境界 6550万年前白亜紀末と新生代

5億4500万年前
V-C境界
ベンド紀ーカンブリア紀の頭文字を取りv-c
原生代末
V-C境界と呼ばれ、最近の研究で大量絶滅があったことが判明しつつある。下に述べる古生代末の大量絶滅(P-T境界)と同じく、超大陸の形成と分裂が原因と推定されている事件。ゴンドワナと呼ばれている超大陸が形成・分裂した時期に相当する。超大陸の分裂に際してはスーパープルームが地上まで上昇してきて非常に大規模な火山活動が起こり、地球表面の環境が激変するため、大量絶滅が起こると考えられている。
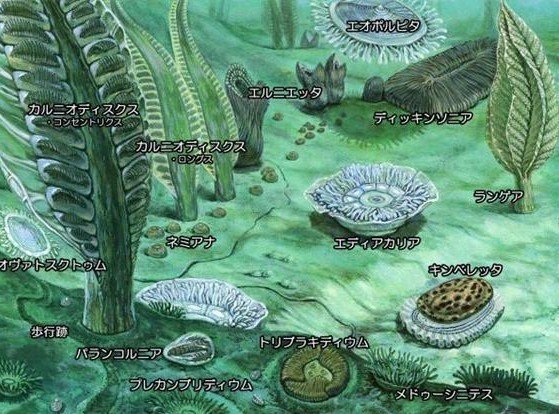
原生代のエディアカラン紀にはエディアカラ生物群[注釈 3]が存在していた。この生物群はオーストラリアのエディアカラで多数の化石が発見されたことから命名された。生物体は全て軟組織でできており、体表を保護する硬い骨格を有していなかった。エディアカラ生物群は約5億4500万年前のV-C境界を境にほとんど見つからなくなるが、以後、三葉虫のような硬骨格を有する生物が出現する。
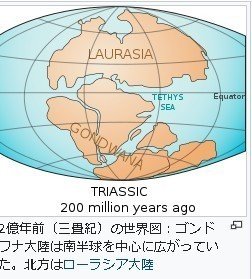
ゴンドワナ大陸は、今から約2億年ほど前から分裂を始め、中生代白亜紀末(6500万年前)にはアフリカから南米、南極、インド、オーストラリアの各プレートが離れたとされている
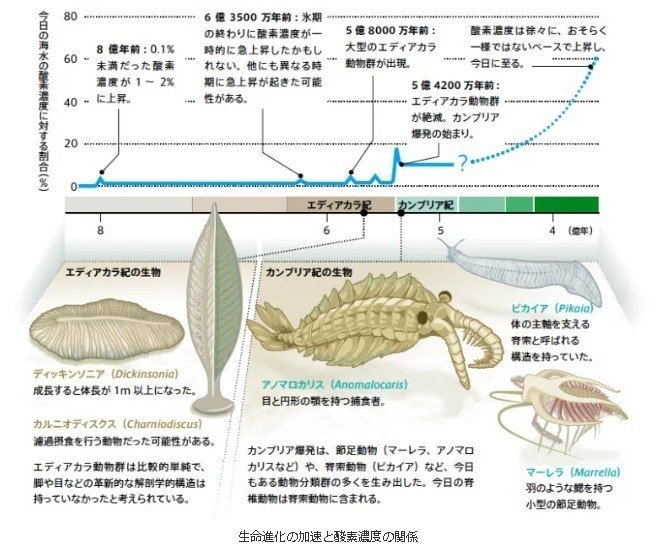
カンブリア紀
5億4100万 - 4億8540万年前
三葉虫、アノマロカリスなど無脊椎動物の繁栄。
生物種の爆発的増加(バージェス動物群)。
最古の脊椎動物である無顎類の登場
カンブリア爆発
酸素濃度上昇とともに急激に種の多様性が生れる
5億4200万年前
葉足動物メガディクティオン
後に節足動物に分岐

5億2,400万年前
古生代カンブリア紀前期中盤[カエルファイ世アトダバニアン]
最初の魚類
ミロクンミンギア誕生
澄江動物群の一つであり、広義で言うところでは既知で最古の魚類である。

5億2,100万年前
初期の節足動物
三葉虫
最初に眼を獲得した生物

オパビニア

カナダスピス

ハルキゲニア

オットイア 鰓曳動物

ヒオリテス オットイアに多数捕食された。

ディアニア 前期カンブリア紀の地層よりただ一種が発見されている。節足動物の起源に非常に近い可能性がある。

ディノミスクス

4億8800万年前
カンブリア紀末
古生代のカンブリア紀末(約4億8800万年前)に大量絶滅が発生し、三葉虫、腕足類、コノドントが激減した。又、カンブリア紀には約5億1700万年前及び約5億200万年前にも大量絶滅がたて続けに起きている。

オルドビス紀
4億8540万 - 4億4380万年前
オゾン層の形成。
フデイシ(筆石)の繁栄。
前の時代に引き続き三葉虫が繁栄した。
オウムガイの繁栄。
4億5000万年前
最初の陸上植物
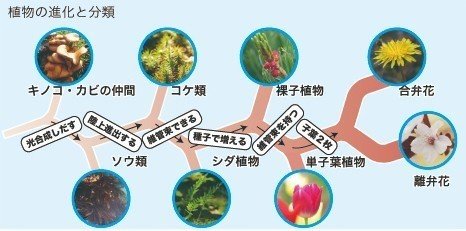
4億4310万年前から4億4290万年前
O-S境界
オルドビス紀末ーシルル紀
古生代のオルドビス紀末(約4億4400万年前)に大量絶滅が発生し、それまで繁栄していた三葉虫、腕足類、ウミリンゴ、サンゴ類、筆石、コノドントの大半が絶滅した。当時生息していた全ての生物種の85%が絶滅したと考えられている。
この時期、大陸は南極域にあり、短い期間であるが大陸氷河が発達した。絶滅は、氷床の発達に伴う海水準の低下時及び氷河の消滅に伴う海水準の上昇時の2回確認されているが、海水準変動をもたらした環境の変化と大量絶滅との関係は不明である。
2005年、アメリカ航空宇宙局(NASA)とカンザス大学の研究者により、近く(6000光年以内)で起こった超新星爆発によるガンマ線バーストを地球が受けたことが大量絶滅の引き金となった、という説が出されている。
2017年、東北大学大学院などの研究チームは、火山噴火による地球寒冷化が原因とする仮説を発表した。
2020年、中国、アメリカ、オーストラリアの研究チームは、オルドビス紀末の大量絶滅が4億4310万年前から4億4290万年前までの20万年の間に発生したとの見解を発表した。これは雲南省永善県で発見されたオルドビス紀とシルル紀の地層が完全に連続している境界面を分析した成果による

可能性のある原因
オルドビス期末の大量絶滅の結果として泥から飛び出したカメロケラスの復元図。
南極に位置していたゴンドワナ大陸の急速な氷床の発達・寒冷化・世界の海洋の全水柱における流体力学的な変動・対応する海水準の低下はストレス状態の前提となっており、オルドビス紀の海洋生物相の主要な生態系ニッチである大陸棚と浅海盆の減少をもたらした。他の大量絶滅現象と同様に、これらの過程には火山活動やそれによる光合成や生物生産性の低下・食物連鎖の破壊・海洋無酸素事変を伴っていた。大気中の二酸化炭素を消費する陸生藻類や微小植物プランクトンの出現と繁栄は、温室効果を減少させて気候システムの氷河期時代への移行を促進するという、この時期に特有の役割を果たした。
氷床の発達
氷河時代に伴う2つの環境変動が後期オルドビス紀の絶滅の大部分の原因となった。1つは当時の生物が強烈な温室効果に適応していたため全球的な気候の寒冷化が特に有害であったことである。もう1つは、氷床の形成で水が集積されて海水準が低下し、広大な内海が失われて数多くの固有の生物の生息地が消失したことである。
ゴンドワナ氷床の存在は顕生代で最も苛烈な氷河時代の始まりと終わりに対応するとみなされており、この氷河期は中期ヒルナンシアン期で終わりを迎えた。
南半球の超大陸ゴンドワナが南極点を超えると、大陸上に氷床が形成された。当時の南極点であった南アメリカ北東部およびそれに隣接する北アフリカの上部オルドビス系からはこれに対応する岩石層が検出されている。世界中の水が氷河に凝固した後、間氷期には水が解放され、海水準が劇的に上下を繰り返した。最初の海退で多くの生息地を失った当初の生物群は海進で戻ってきた後、さらなる海退を受けて激減を続けた。北アフリカでは、地震断面から氷河の5回のパルスが記録されている。
ガンマ線バースト
最初の絶滅は地球から6000光年以内(銀河系の太陽系近くの腕)の超新星に由来するガンマ線バーストで引き起こされたと提唱した科学者もいる。10秒のガンマ線バーストで地球大気のオゾンの半分が即座に剥ぎ取られ、惑星の光合成に携わる生物を含む地表生物の多くが高レベルの極端紫外線放射を浴びただろうと推測された。この仮説は絶滅の始まりのパターンと一致するが、このようなガンマ線バーストが地球の近くで起こったという証拠はない。
火成活動
後期オルドビス紀の氷河作用(英語版)に先駆けて、大気の二酸化炭素濃度が 7,000 ppm から 4,400 ppm へ低下していた。火山活動により堆積した新しい珪酸塩岩は浸食をうけると大気中に二酸化炭素が引き出される。二酸化炭素の重要な役割は2009年の論文で示されている。大気と海洋の二酸化炭素レベルはゴンドワナ氷床の成長・崩壊に伴って変動した可能性がある[11]後期オルドビス紀を通して、大規模な火山活動からのガス放出はアパラチア山脈の隆起による激しい風化によってバランスがとれていた。ヒルナンシアン期になると火山活動は停止し、継続的な風化によって二酸化炭素は急激に減少することとなった[21]。これは急速で短い氷河期と一致する。
また、火成活動は必ずしも温暖化に寄与したとは限らない。火山から噴出した二酸化硫黄が大気中で硫酸のエアロゾルを生み出し、それが地表に届く太陽光を遮蔽して気候の寒冷化、そして氷床の発達に寄与したとも考えられている。また、同研究では以下に説明する有害金属も火成活動に由来すると推論されている。
金属汚染
海洋の酸素が減少したことにより鉛やヒ素や鉄といった海底の有害金属が水中に溶け込んだ可能性も指摘されている。生物が利用可能な海洋の栄養素の増加、また、地球規模の冷却による海洋循環の低下が要因となっている可能性がある。有毒金属は食物連鎖の下位の栄養段階にある生物を死に至らしめ、個体数の減少を引き起こし、その結果として食物連鎖の上位に位置する摂食生物の餓死をもたらした可能性がある。現在のリビア砂漠に分布する当時の地層では、プランクトンの奇形が通常の100倍以上の頻度で確認できる。
これらの金属は先述する火成活動に由来する可能性もある。日本の東北大学などの研究では、中華人民共和国とアメリカ合衆国に分布するO-S境界付近の地層から高濃度の水銀が検出されており、大規模な火成活動で水銀が大気中に放出されたと推測されている。
絶滅事変の終わり
氷床が融解して海水面が上昇すると再び絶滅のピークが訪れたが、それが安定すると絶滅事変も収束を見せた。シルル紀の初期に大陸棚が長期間再び冠水したため、生命の多様性は回復し、生き残った生物群集の中で生物多様性が増加した。
このように生物多様性が大きく失われた後、シルル紀の生物群集は最初に、より複雑ではなく、より広範囲に分布するようになった。後期オルドビス紀の特徴であった固有性の高い動物群集は、顕生代で最も世界的な動物群集に取って代わられ、シルル紀のほとんどの期間にわたって持続する生物地理学的パターンを呈した。
オルドビス紀 - シルル紀の大量絶滅事変はペルム紀末の大量絶滅や白亜紀末の大量絶滅のように長期に亘る爪痕を残したわけではなかったが、短期間のうちに数多くの分類群が地球上から姿を消し、多様性は失われて変化していった。
4億7900万年前
オルドビス紀
甲殻類から派生した六脚類が陸上に生活圏を移動
4億4千万年前
シルル紀
4億4380万 - 4億1920万年前
顎やウロコを持つ魚類の登場。
サンゴ類の繁栄(ファボシテスなど)。
植物の陸上進出(クックソニアなど)。
昆虫の誕生。
脊椎動物が一般的になった。
最初の昆虫は、現在のカマアシムシ、トビムシに近い無翅昆虫であった。 間もなく昆虫は翅を手に入れる。 昆虫類の系統関係から有翅昆虫の起源を調べた結果、無翅昆虫の中でもシミ目が有翅昆虫に近縁であることがわかった。 この結果は、シミ目が有翅昆虫と形態的にも近縁であることと一致する。
デボン紀
4億1920万 - 3億5890万年前
海域を中心に板皮類が繁栄(ダンクレオステウス、ボトリオレピスなど)。
淡水域を中心に棘魚類が栄えた。
硬骨魚類が増加した。
オウムガイ、アンモナイトの繁栄。
ハイギョの出現。
種子植物の出現。
最初の森林の形成(アーケオプテリスなど)。
シダ植物の繁栄が始まる。
動物の陸上進出。
両生類の出現(アカンソステガ、イクチオステガなど)。
後期には大量絶滅があった。
魚類の種類や進化の豊かさと、出現する化石の量の多さから、「魚の時代」とも呼ばれている。
4億600万年前
最初の翅で飛ぶ昆虫 有翅昆虫誕生
3億7400万年前
F-F境界
デボン紀後期ー石炭紀初期
特にサンゴ礁を作る赤道域の浅海域で選択的に絶滅が起こっている。この大絶滅により、海洋生物種の82%が絶滅した。その中には、デボン紀に繁栄を極めたダンクルオステウスなどの板皮類や、原始的な脊椎動物である無顎類の大部分や、プロエタス目を除いた三葉虫の大部分が含まれる。
気候の急激な寒暖の変化、海水面の後退、乾燥化、低酸素化、などの大きな環境変化がデボン紀後期に繰返し発生し、おそらくこれらの環境変化が大量絶滅の要因だとは考えられている
腕足類や魚類のデータから、高緯度より低緯度の、淡水域より海水域において絶滅率が高いことが判明している。
この時期の環境の変化として、寒冷化と海洋無酸素事変の発生が知られている。酸素及び炭素同位体比のデータは、2度の寒冷化及び有機物の堆積及び大気中の二酸化炭素の減少を示しており、これは、海水準の上昇及び大量絶滅と同時に起こっている。また、海水中のストロンチウム同位体比の変動は、大陸風化の増加(気温の上昇)を示している。ベルギー及び中国南部のF/F境界層から、小天体衝突の証拠となるスフェルールが報告されているものの、大量絶滅との関連はわかっていない
ダンクルオステウス(板皮類)がここで全滅

原因
この出来事はケルワッサー事変とも呼ばれ、F-F境界とケルワッサー海洋無酸素事変層は一致する
仮説
巨大隕石衝突
シリヤン・リング、スウェーデンにあるクレーター3億7680万年前
3億6000万年前
デボン紀
最初の両生類。イクチオステガ
形はイモリに似ているが、全長1mと巨大である。

石炭紀
3億5920万年前ー2億9900万年前まで
シダ植物の繁栄。
板皮類の絶滅。
両生類の地上上陸。
有羊膜類(爬虫類と哺乳類の共通祖先)の出現。
巨大昆虫類の繁栄(パレオディクティオプテラなど)。
古生代の後半で、デボン紀の後、ペルム紀の前の時代を指し、この期間はデボン紀末の大量絶滅からペルム紀直前の数百万年に及ぶ氷河時代で区切られている。
名前の由来はこの時代の地層から多く石炭を産することによる。この地層から石炭を産するのは当時非常に大きな森林が形成されていたことの傍証となる。

3億年前、石炭紀の後期に二酸化炭素濃度は現代の程度まで低下する。この前後、寒冷化が起きた。一方で酸素濃度は地球史上最高の35%となっていた。これは植物の活動(光合成)が大きい。当時、リグニンを含む樹木は腐敗分解されず、石炭化していくのみであった。しかし、これ以降、樹木を分解できる菌類(白色腐朽菌)が登場し酸素濃度は徐々に減少に向い、逆に二酸化炭素濃度は増加に向かう。白色腐朽菌は倒木を分解でき、これにより石炭紀が終焉し、さらに大量の倒木の分解により酸素を大量に消費し二酸化炭素を増大させていった。ペルム紀を通じてこの傾向は続き、P-T境界で以降の地球の低酸素環境は決定的となった。

古生代石炭紀の森林に生息した超巨大ヤスデの1属「アースロプレウラ」。全長2.4メートルほどにも成長する
3億5千万年前
最初の完全変態昆虫
3億1,500万年前
カナダで最初の爬虫類ヒロノムス誕生
爬虫類は、古生代の石炭紀(3億5900万年前~2億9900万年前)に登場し、中生代(2億5100万年前~6550万年前)に大繁栄した
ペルム紀
2億9890万 - 2億5190万年前

棘魚類の絶滅。
両生類・爬虫類・単弓類(哺乳類型爬虫類)の繁栄。
三葉虫類の絶滅。
巨大大陸パンゲアの出現。
ペルム紀末(P-T境界と呼ばれる古生代と中生代の境界)では、史上最大とされる地球規模の大量絶滅が起こった。

メガネウラ(2億9000万年前ー2億5,100万年前)
全長は70cm、当時の地球は酸素濃度が高かった(32%)ため、昆虫たちは軒並み巨大化。メガネウラも進化して巨大になった。濃い酸素が巨体の維持を可能としていた。メガネウラには肺が備わっており、酸素を効率よく活用して大空を駆け巡っていた。
ペルム紀末期の酸素濃度低下に適応できず絶滅
2億5100万年前
P-T境界
ペルム紀末ートリアス紀(三畳紀)
地球の歴史上最大の大量絶滅が起こった。海生生物のうち最大96%、全ての生物種で見ても90%から95%が絶滅した。既に絶滅に近い状態まで数を減らしていた三葉虫はこの時に、とどめをさされる形で絶滅した。
2億5100万年前に位置する谷間がP-T境界

原因
超大陸の形成 大陸が1つ、海洋が1つになって多様性が減少
シベリア洪水玄武岩 過去6億年間でもっとも大きな火山噴火
スーパーアノキシア 何らかの原因で海洋が低酸素化した
海水準の変化 寒冷化で急激な海水準の低下
炭素同位体比の急変 12Cと質量数13の13Cの変化についていけない
巨大隕石の落下の可能性 まだ見つかっていない
上ほど有力
急速な大陸移動により海溝より深くにマグマが潜り込むエネルギーが発生

地球の核まで達し、潜り込むエネルギー分、核から地表にマグマが移動(スーパープルーム)

過去にないマグマが地表に出る
地球温暖化

温暖化で深海のメタンハイドレートが溶ける

メタンの温室効果によりさらにメタンハイドレートが溶ける温暖化悪循環
ペルム紀の末期には激しい気温上昇が起こり、地球の平均気温は23℃にも達した。これは、6億年前から現在まででもっとも高い気温である。
この大量絶滅によりペルム紀は終わり、中生代三畳紀へ
三畳紀
2億5190万年前ー2億130万年前

恐竜の時代が始まる
古生代末、ほとんど全ての大陸が合体し、三畳紀には北極から南極に至るパンゲア大陸と呼ばれる超大陸が形成された
また、山地をくずして内陸部に広大な平野をつくる陸地の平原化現象がおおいに進行した。
内陸部の平野には乾燥気候の影響で砂漠化の進行がいちじるしく、赤色の砂が堆積していった。砂漠のところどころにはオアシスが点在した。
古生代終期に寒冷化した気候も、三畳紀を通じて気温は徐々に上昇していったものと推定される。ペルム紀に30パーセントほどあった酸素濃度も10パーセント程度まで低下し、ジュラ紀頃までの約1億年間、低酸素状態が続いた。
三畳紀は、広大な大テチス地向斜の発展がみられた時期と考えられている。この地向斜から、2億もの年月を経たのち、アルプス・ヒマラヤ造山帯など新期造山帯と称される若い山脈が形成されていくものとみられている
2億5000万年前から6500万年前
トリケラトプス

2億2,500万年前
中生代三畳紀後期
最初の哺乳類
繁殖は卵を産んでいた。
それまで最古の哺乳類であった2億1,000万年前のエオゾストロドンより、さらに1,500万年さかのぼることになった。

2億1,500万年前
T-J境界
三畳紀(トリアス紀)ージュラ紀
T-J境界と一致する衝突クレーターは発見されていない
三畳紀の終わりに、再びやや小規模な大量絶滅があった。海洋ではアンモナイトの多くの種が姿を消し、魚竜などの海洋棲爬虫類も打撃を受けた。陸上ではキノドン類、ディキノドン類の大半の種といった大量の単弓類(哺乳類型爬虫類)が絶滅した。三畳紀の終末を生き延びた恐竜など陸生脊椎動物は、繁殖様式(卵など)や生活様式から乾燥にとくに強いタイプのものと考えられる。また、爬虫類も単弓類同様に大型動物を中心に多くの種が絶滅した。まだ比較的小型だった恐竜は、三畳紀末期には竜脚類のような大型種も出現し、そののち急速に発展していく。絶滅の原因としては、直径3.3 - 7.8km程度の隕石の落下あるいは、中央大西洋マグマ分布域における火山活動との関連が指摘されている。

ここで絶滅した単弓類 ディメトロドン(体長約3m)

三畳紀末の大量絶滅、原因は溶岩の噴出?
三畳紀の終わりに、地球上の陸と海の生物の少なくとも半分が絶滅した。これを契機に恐竜が勢力を拡大し、その後1億3500万年にわたり地球を支配することになる。専門家の間では長らく、この大量絶滅の原因は大規模な火山の噴火と考えられていたが、これまでその正確な年代は特定できていなかった。 このほど発表された研究によると、多数の動植物種が化石にその痕跡を残さなくなるのと同時期に、非常に大規模な溶岩の噴出が起こっていることが確認された。この溶岩がアメリカ一国の上に堆積したと仮定すると、その高さは90メートルを超える計算になる。
今回の研究の主著者で、ワシントンD.C.にあるカーネギー研究所で地質学を研究するテレンス・ブラックバーン(Terrence Blackburn)氏は次のように説明してくれた。
2億100万年ほど前に、超大陸パンゲアがプレートの運動によって分裂を始めたが、このとき、地殻の下にあるマントルの岩石が溶けて、これほど大きな噴出につながったというのだ。
このとき噴出した溶岩は洪水玄武岩と呼ばれる。60万年の間に4回に分けて噴出したが、そのうち最初の火山活動が、多くの生物種の絶滅につながったとブラックバーン氏は見ている。
古代のワニの祖先や、コノドントというウナギのような生物、哺乳類型爬虫類とも呼ばれる獣弓類など多数の生物種が、このとき根絶したと見られる。というのも、この時期を境に化石として残らなくなっているからだ。
◆ジルコンはタイムカプセル
これまで、この溶岩の噴出の時期には100~300万年の誤差が想定されていた。そのため、溶岩の噴出と大量絶滅のどちらが先に起きたのか、はっきりしていなかった。
ところが今回の研究では、玄武岩などの火成岩に含まれる希少鉱物のジルコンを使って、この誤差範囲を2~3万年までに縮めることができた。これによって、最初の噴出は大量絶滅の直前に起こったものと特定できた。
「ジルコンはこれらの岩石の年代を特定する上で完璧なタイムカプセルだ」とブラックバーン氏は言う。ジルコンは結晶化の際にウランを取り込みやすい。そしてウランの原子核は一定の期間で崩壊し、鉛の原子核に変化する。それゆえ、試料に含まれるウランと鉛の比率を測定することで、その結晶の年代を特定できるとブラックバーン氏は説明する。
大量絶滅が溶岩の噴出の最初の段階で起こったと正確に確認できたことで、今後は気候モデルなどを専門とする研究者らによって、この絶滅を引き起こした具体的な要因(たとえば気候変動など)が特定されるのではないかとブラックバーン氏は言う。
溶岩の噴出の最初の段階で大気中の二酸化炭素量が2倍になり、地球規模の気温上昇や海水の酸性化につながったことを示す証拠は見つかっている、とブラックバーン氏は言う。これらは急速に起こったため、ほとんどの生物はその変化に適応しきれなかった。
ただし、2度目以降の溶岩噴出の際には生命の復活のきざしが見られるとブラックバーン氏は言う。これはブラックバーン氏にとって驚くべきことだったそうだ。
たしかに、2度目以降に噴出した溶岩の量は、初回ほどは多くない。しかし別の研究グループによれば、大気中の二酸化炭素量はやはり噴出のたびに増加しており、大量絶滅の際の気候条件がその後も続いていた可能性が指摘されている。
大量絶滅を生き延びたと見られる生物種が、いかにして新しい気候に適応できたのかは、いまだ不明である。
◆隕石衝突説の陰で
大量絶滅は歴史上何度も起こっており、そのすべてに大規模な火山の噴火が関係していると、カリフォルニアにあるバークレー地質年代学センターで岩石の年代特定を専門にするポール・レニ氏は言う。しかし、火山活動が主な生物種を一掃するほどの役割を果たしたとする説は、これまであまり検討されておらず、大量絶滅には隕石の衝突が関係しているとする仮説のほうが好まれてきた。
たとえば6500万年前の白亜紀/第三紀境界(K-Pg境界)と呼ばれる時期には恐竜が大量絶滅しているが、これとほぼ同時期に隕石の衝突があったことを裏付けるたしかな証拠が見つかっている。
「この1つの事例が皆の考え方を方向づけてしまい、隕石の衝突が大量絶滅につながったという見方がすっかり一般的になってしまった」とレニ氏は言う。レニ氏は今回の研究には参加していない。
しかしレニ氏は、大規模な火山の噴火こそが大量絶滅の原動力であったと見ており、ブラックバーン氏らの最新の研究によって、三畳紀末に起こった溶岩の噴出と大量絶滅という2つの出来事を結びつけて考える根拠が強まったと見ている。
レニ氏は、K-Pg境界の大量絶滅に際しても、インドで始まった大規模な火山の噴火が主要な役割を果たしており、隕石の衝突はそこへとどめの一撃を加えたにすぎないという可能性を指摘している。

海ではコノドントが絶滅し、陸上ではワニ形上目とアヴェメタターサリア(翼竜と恐竜)を除く全ての主竜類、生き残りの獣弓類、数多くの大型両生類が絶滅した。この時代における海洋生命の消失の統計的解析によると、多様性の消失は絶滅の増加よりも種分化の減少によるところが大きいことが示唆されている。
原因や仮説
隕石衝突 可能性は低い

火山の噴火 噴火による寒冷化など
緩やかな過程 植生の変化、酸素の減少など複合的要因で緩やかに大量絶滅
セコイア

セコイアは、ヒノキ科(またはスギ科)セコイア属の常緑針葉樹。セコイア属はセコイアのみの1属1種である。
高さ100m近くにもなる世界有数の大高木。アメリカ合衆国西海岸の沿岸一帯に自生する。
セコイアスギ、センペルセコイア、レッドウッド、アメリカスギ、あるいはコースト・レッドウッド、カリフォルニア・レッドウッドなどとも呼ばれる。セコイアデンドロンとの対比からセコイアメスギとも、葉の形が似ている事からイチイモドキとも呼ばれる。
世界一の樹高を誇り、カリフォルニア州レッドウッド国立州立公園のセコイアは樹高世界1位から3位までを独占する。平均的な大きさは樹高80メートル、胸高直径5メートル、樹齢は400年から1300年ほどで、2200年のものが現在知られる最高齢である。厚さ30センチに及ぶ樹皮や心材の色からレッドウッドとも呼ばれる。この樹皮と木質部はタンニンを多く含み、病原菌や白蟻の侵入を拒む。
この厚い樹皮は、他の広葉樹が燃え尽きてしまうような山火事の際にも木の内部を守る。北アメリカ西海岸に見られるセコイアの純林は、度重なる山火事によりできあがったものと考えられている
ジュラ紀
1億9960万年前ー1億4550万年前まで

ジュラ紀の開始は三畳紀末の大量絶滅から始まった。
最後の巨大な両生類もこのときにほぼ姿を消した。
三畳紀末期の絶滅を生き残った恐竜が栄えた。
三畳紀から相次いだ火山活動の結果、大気中の二酸化炭素濃度は高く、ジュラ紀は現在よりも暖かく、降水量も多く、湿度も高かった。そのため動物、植物はともに種類が増え、大型化していった。植物ではイチョウ、ソテツなどの裸子植物が大きく繁栄し、それまで植物が無かった内陸部まで生育範囲を広げていった。またジュラ紀の後半には被子植物も現れた。海洋ではアンモナイトや、プランクトンが繁栄し、地上では恐竜が多種多様な進化を遂げた。小型の恐竜の一部が鳥類に至る進化を果たし、始祖鳥が現れたのもこの時代である。
ジュラ紀にもっとも進化した生命は海洋での魚類と、海洋で暮らす爬虫類(魚竜、首長竜など)である。また無脊椎動物にはいくつかの新しいグループが現れた。
1億6100万年~1億5100万年前
ジュラ紀後期
最初の鳥類の近縁
トロオドン
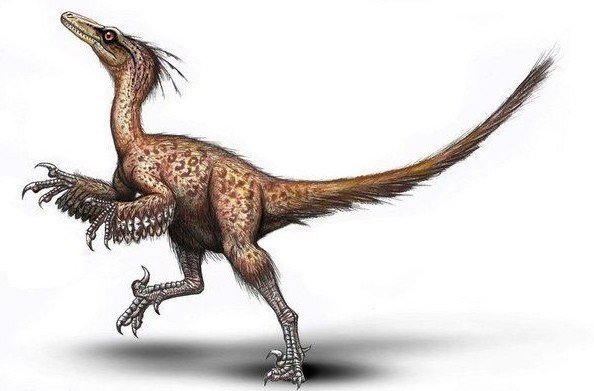
始祖鳥よりも真鳥類に近縁で、原始的な鳥類と見なされることもある。
アーケオプテリクスは現生の鳥類の祖先に近い生物であるものの、直接の祖先では無いと考えられている。
1億5,500万 - 1億5,000万年前
中生代ジュラ紀後期
大型肉食獣脚類
アロサウルス

1億5,500万 ~ 約1億5,000万年前
ジュラ紀後期
剣竜類
ステゴサウルス

1億4000万年前
ジュラ紀
最初の被子植物アルカエフルクトゥス

白亜紀
1億4,500万年前から6,600万年前

温暖な気候と高海水準とで特徴付けられる時代である。
ジュラ紀から白亜紀の境目に大きな絶滅などはなく、長期にわたり温暖で湿潤な気候が続いた。前期白亜紀において、一時的な寒冷化が見られるものの、同時期の表層海水温に関する研究では、低緯度地域で32 ℃、中緯度地域で26 ℃と現在より高い海水温で安定していたことがわかっている。末期には気候帯が現われ、植物相にも変化が見られた。
植物は、主流であった原始的な裸子植物やシダなどが減少し、被子植物が主流となって進化、繁栄を遂げた。スギなどの針葉樹は現代と同じ形まで進化し、イチジク、スズカケノキ、モクレンなどが現在とほぼ同じ形となった。
超大陸パンゲアの分裂が一層進んだ。
これによって地理的な隔離が起きたため、陸上の生物の多様性を更に高めることとなった。
1億1,200万前 - 9,350万年前
中生代白亜紀前期~後期
スピノサウルス

8,930万 ~ 7,400万年前
中生代白亜紀後期
翼竜
プテラノドン

約8500万年前
白亜紀後期サントン階
首長竜
フタバスズキリュウ
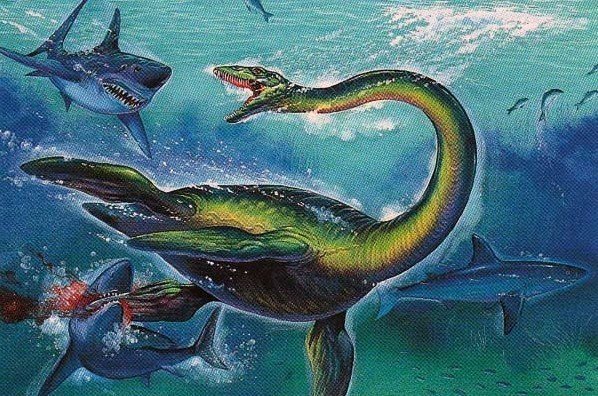
7,000万年前 - 6,600万年前
水生有鱗目
モササウルス

6,800万年前
中生代白亜紀末期
ティラノサウルス

6550万年前
K-Pg境界
白亜紀末
巨大隕石による恐竜絶滅
チクシュルーブ・クレーター

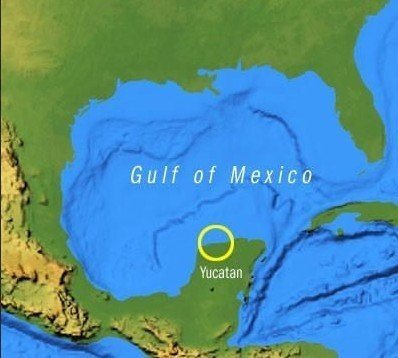


ハスやスイレンの化石から、隕石は6月頃に落下したこと、落下直後には植物が凍結したことが分かった
陸上生物の約50%、海洋生物の約75%、生物全体で約70%が絶滅した。
600万年前ー130万年前
猿人がアフリカ大陸に出現、初期の人類

猿人は脳の重さが3分の1なのでまだ現代の人とは明確に違う
この頃から2足歩行し手で道具を使うようになる
240万年前
原人登場
北京原人、ジャワ原人
石器を鋭利にしたり、洞窟に暮らしたりするようになる。北京原人は火も使用する。

40万年前ー4万年前
ネアンデルタール人(旧人)
旧石器時代

ネアンデルタール人は、ヨーロッパ大陸を中心に西アジアから中央アジアにまで分布しており、旧石器時代の石器の作製技術を有し、火を積極的に使用していた。
ネアンデルタール人類は現生人類と類人猿との中間の特徴を持ち、曲がった下肢と前かがみの姿勢で歩く原始的な人類(原始人)とした。
ネアンデルタール人の脳容量は現生人類より大きく、男性の平均が1600 cm3あった(現代人男性の平均は1450 cm3)。しかし、頭蓋骨の形状は異なる。脳頭蓋は上下につぶれた形状をし、前後に長く、額は後方に向かって傾斜している。また、後頭部に特徴的な膨らみ(ネアンデルタール人のシニョン)がある。
彼らの文化はムステリアン文化と呼ばれ、旧石器時代に属している。
20万年前
新人登場
クロマニヨン人
現代人とほぼ同じ姿かたちをしていた。代表はクロマニヨン人。狩猟採集時代。

紀元前3500年前ー紀元前4世紀
メソポタミア文明

バビロン
チグリス・ユーフラテス両河は水源地帯の雪解けにより定期的に増水するため、運河を整備することで豊かな農業収穫が得られた。初期の開拓地や文化から始まり、エジプトなどよりも早く農業が行われた地域として知られている。紀元前3500年前ごろにメソポタミア文明がつくられた。
ジッグラトと呼ばれる階段型ピラミッド(聖塔といわれているが詳細は不明)を中心に、巨大な都市国家を展開した。また、農耕の面でも肥沃な大地・整備された灌漑施設・高度な農耕器具により単位面積当たりの収穫量は現代と比較しても見劣りしなかったという。さらに、旧約聖書との関連も指摘されており、始祖アブラハムはメソポタミアの都市ウルの出自とされている。エデンの園はメソポタミアの都市を、バベルの塔はジッグラトを、ノアの洪水は当地で突発的に起こる洪水を元にした逸話との説がある。
暦は太陰太陽暦
文字は象形文字を発展させた楔形文字
記録媒体は粘土板
ハンムラビ法典
焼き畑農業の起源
初期メソポタミアでは、南部のシュメール人たちは言語系統不明のシュメール語を、北部のアッカド人たちはセム語族のアッカド語を使用していた。
北部メソポタミアでは、後期新石器時代に入ると紀元前6000年から紀元前5500年ごろのハッスーナ期
紀元前5600年ごろから紀元前5000年ごろにかけてのサーマッラー期
紀元前5500年ごろから紀元前4300年ごろにかけてのハラフ期の、3つの文化が栄えていた
メソポタミア(ギリシャ語: Μεσοποταμία、ラテン文字転写: Mesopotamia、ギリシャ語で「複数の河の間」)は、チグリス川とユーフラテス川の間の沖積平野である。現在のイラクの一部にあたる。
古代メソポタミア文明は、メソポタミアに生まれた複数の文明を総称する呼び名で、世界最古の文明であるとされてきた。文明初期の中心となったのは民族系統が不明のシュメール人である。
地域的に、北部がアッシリア、南部がバビロニアで、バビロニアのうち北部バビロニアがアッカド、下流地域の南部バビロニアがシュメールとさらに分けられる。南部の下流域であるシュメールから、上流の北部に向かって文明が広がっていった。土地が非常に肥沃で、数々の勢力の基盤となったが、森林伐採の過多などで、上流の塩気の強い土が流れてくるようになり、農地として使えない砂漠化が起きた。
古代メソポタミアは、多くの民族の興亡の歴史である。 例えば、シュメール、バビロニア(首都バビロン)、アッシリア、アッカド(ムロデ王国の四つの都市のひとつ)、ヒッタイト、ミタンニ、エラム、古代ペルシャ人の国々があった。
古代メソポタミア文明は、紀元前4世紀、アレクサンドロス3世(大王)の遠征によってその終息をむかえヘレニズムの世界の一部となる。

紀元前3000年ー紀元前30年
エジプト文明

古代エジプトは、古代のエジプトに対する呼称。具体的に、どの時期を指すかについては様々な説が存在するが、この項においては紀元前3000年頃に始まった第1王朝から
紀元前30年にプトレマイオス朝が共和制ローマによって滅ぼされるまでの時代を扱う。

紀元前2600年ー紀元前1800年
インダス文明

インダス文明(インダスぶんめい、Indus Valley civilization)は、インド・パキスタン・アフガニスタンのインダス川および並行して流れていたとされるガッガル・ハークラー川周辺に栄えた文明である。
これら各国の先史文明でもある(インドの歴史、パキスタンの歴史、アフガニスタンの歴史も参照)。
崩壊の原因となったという説のあった川の名前にちなんでインダス文明、最初に発見された遺跡にちなんでハラッパー文明とも呼ばれる[1]。
狭義のインダス文明は、紀元前2600年から紀元前1800年の間を指す。インダス文明の遺跡は、東西1500km、南北1800kmに分布し、遺跡の数は約2600におよぶ。
そのうち発掘調査が行われた遺跡は、2010年時点でインド96、パキスタン47、アフガニスタン4の合計147となっている
言語
インダス文明の言語は原ドラヴィダ語に属すると推定されている。
文字
インダス文字は現在でも未解読である。統計的分析ができる長文や、ロゼッタ・ストーンのように多言語併記の物が出土しないことが研究の大きな障壁になっている。一方で、インダス式紋章は文字ではないという説もあり、論争が続いている
牧畜
現在でも家畜として飼育されているコブウシは、インダス文明の土器の模様、印章、土偶などのモチーフにも多数使われている。コブウシよりは少ないがコブのないウシも描かれており、系統の異なるウシが飼育されていた可能性がある
鉄は知られず、青銅器を使った。都市計画で知られるように建築技術に優れており、建築物には縦:横:厚みの比が4:2:1で統一された焼成煉瓦が広く使われている。服は染色された綿で作られていたようで、染色工房と推定される場所が見つかっている。

インダス文明の衰退や滅亡については次のような諸説がある。
砂漠化説
インダス文明が存在した地域は現在砂漠となっている。インダス文明が消えたのは、この砂漠化によるのではないかという説がある。
砂漠化の原因としては、紀元前2000年前後に起こった気候変動があげられている。
大西洋に広がる低気圧帯は、一時北アフリカと同じ緯度まで南下し、さらにアラビア・ペルシア・インドにまで及んで、雨をもたらし、緑豊かな土地になっていた。
しかしやがてこの低気圧帯は北上し、インドに雨をもたらしていた南西の季節風も東へ移動して、インダス文明の栄えていた土地を現在のような乾燥地帯にしてしまった、という説である。
衰退後の植物相や動物相には大きな変化が見受けられないことから、気候の変動を重視する説は見直されている。
インダス文明が森林を乱伐したために砂漠化が進行したという説もある。
しかし、乾燥化説については、ラクダの骨や乾地性のカタツムリが出土していること、綿の生産が行われていたことなどは、川さえあれば気温の高い乾燥ないし半乾燥地帯で文明が興りえたことを示し、「排水溝」も25ミリの雨がふっただけでももたない構造であり、煉瓦を焼くにも現在遺跡の周辺で茂っている成長の早いタマリスクなどの潅木でも充分間に合ったのではないかという反論があり、決定的な説となってはいない。
河流変化説
紀元前2000年頃に地殻変動が起こり、インダス川の流路が移動したために河川交通に決定的なダメージを与えたのではないかという説。
インダス遺跡はインダス川旧河道のガッカル=ハークラー涸河床沿いに分布している。
気候変動説
気候変動によってインダス文明が衰退したとする説である。
4200年前には、地中海から西アジアにかけて冬モンスーンが弱く乾燥化が起き、メソポタミアではアッカド王国崩壊の一因になったという説がある。
こうしたモンスーン変動がインダス文明の地域にも影響を与えたとされる。
2012年にはアバディーン大学が中心の研究グループが発表し、2013年には京都大学が中心のグループがネパールのララ湖(英語版)を調査して3900年前から3700年前にかけて夏モンスーンが激化していたことを明らかにした[3]。
また、遺跡の数はインダス文明の盛期ハラッパー文化期よりも後期ハラッパー文化期のほうが多く、規模が縮小している。 これらの点から、夏モンスーンの激化がインダス川流域に洪水を起こし、インダス川流域に位置するモヘンジョダロなどの大都市から周辺への移住が起きたとする[4]。
また、インダス文明期には、海面が現在よりも2mほど高かったという調査がある。これにより遺跡の分布を調べると、インダス川流域以外のグジャラートやマクラーン海岸の遺跡の多くが海岸線に近くなる。
そこで、海岸線に近いインダス文明の人々は大河によって生活するのではなく、海上交易などを行っていた海洋民であったが、海面低下により生活が変化したとする説も提唱されている。後述のように、インダス文明はメソポタミアやペルシア湾地域と交易を行っていたことが確認されている。
アーリア人侵入説
「アーリア人」および「アーリアン学説」も参照
インダス文明滅亡の原因は古くから論争があり、第二次大戦後にはM.ウィーラー(英語版)によるアーリア人侵略説をはじめとする外部からの侵略説が唱えられた。
発掘調査によって埋葬もされずに折り重なるおびただしい人骨が確認されたために外部からの侵入による虐殺説が唱えられた。
また、『リグ・ヴェーダ』などの戦争記事がその根拠のひとつとされた。しかし、当時の発掘調査は、層位関係を考えずに地表からの深さのみを記録して行われた調査であったために同時期の人骨ではなかった。
その他、虐殺跡とされた人骨には外傷の形跡がなく、アーリア人の侵入とインダス文明衰退の年代には相違があり、『リグ・ヴェーダ』の記述の史実性にも問題が指摘され、現在では否定されている。
日本においても第2次世界大戦前にアーリアン学説を補強する学説が発表された。
この説では、インダス文明は南インドを中心に暮らしているドラヴィダ人の祖先によりつくられたと推定されている。また、ドラヴィダ人は、紀元前13世紀に起きたアーリア人の侵入によって、被支配民族となり先住民族であるドラヴィダ族を滅ぼしてヴァルナという身分制度を作り上げたという説がある。
紀元前5000年-紀元前3000年
黄河文明
黄河文明(こうがぶんめい)は黄河の中・下流域で栄えた古代の中国文明のひとつである。黄河の氾濫原で農業を開始し、やがて黄河の治水や灌漑を通じて政治権力の強化や都市の発達などを成し遂げていった。東アジアの歴史の教科書には「世界四大文明」のうちのひとつとして挙げられていることが多いが、現在は長江文明や遼河文明などさまざまな文明が中国各地で発見されているため、四大文明に黄河文明のみを取り上げる手法はもはや古くなっている(最近の教科書では、「黄河・長江流域の文明」のようにややぼかした表現になっている)。
仰韶文化

次回やりたい事
シュメール人やウイグル人など、どう生まれてどう移動しどうなったかをまとめたいな(っ♥・ ◡・c)
購入されたら他の記事購入資金に使わせてもらいます