非経口BCGワクチンは、腸-肺軸を介して、肺に常駐するメモリーマクロファージと訓練された免疫力を誘導する

公開日:2022年12月01日
非経口BCGワクチンは、腸-肺軸を介して、肺に常駐するメモリーマクロファージと訓練された免疫力を誘導する
Mangalakumari Jeyanathan, Maryam Vaseghi-Shanjani, ...Zhou Xing 著者を表示する。
Nature Immunology volume 23, pages1687-1702 (2022)この記事を引用する

6059 アクセス数
1 被引用数
33 Altmetric
指標詳細
概要
非経口ワクチンや感染症によって骨髄や末梢血に誘導される中枢性訓練免疫とは別に、粘膜曝露後に局所炎症経路を介して粘膜常在性自然免疫記憶が発達することを示す証拠がある。しかし、非経口ワクチン接種/感染後に遠位で生成された免疫学的シグナルを統合することによって粘膜常在型自然免疫記憶が生じるかどうかは不明である。本論文では、Bacillus Calmette-Guérin(BCG)皮下接種が肺胞マクロファージ(AM)を記憶させ、肺で訓練された免疫を誘導することを示す。BCGの非経口接種は肺胞の前駆細胞や循環単球を訓練するが、メモリーAMsの誘導は循環単球とは無関係である。むしろ、非経口BCG接種は、マイコバクテリアの播種を介して、腸内マイクロバイオーム、バリア機能および微生物代謝産物に時間依存的変化をもたらし、その後、循環および肺代謝産物に変化をもたらし、肺におけるメモリーマクロファージの誘導および訓練された免疫につながることが明らかになった。これらのデータは、遠位粘膜組織における自然免疫記憶発達の腸内細菌叢を介した経路を特定し、呼吸器系病原体に対する次世代ワクチン戦略の開発に示唆を与えるものである。
主な内容
宿主防御やワクチン学において、自然免疫記憶や訓練された自然免疫(TII)の重要性が認識されつつある1,2,3。疫学的研究により、弱毒生ワクチン(Bacillus Calmette-Guérin(BCG)を含む)の非経口免疫により、標的および非関連病原体の両方から保護されることが示されている4。したがって、抗結核BCGワクチン接種は、幼児および高齢者の全死亡率および/または呼吸器感染症を減少させます5,6,7。このような全身性/親微生物性、炎症性、あるいはワクチンによるTIIは、主に骨髄(BM)中の骨髄系前駆細胞の代謝/エピジェネティックな再配列の結果、単球を含む中枢で鍛えられた循環白血球を介して行われる8,9,10,11,12,13。
これまで、粘膜に存在するマクロファージが、自然記憶を持続的に保存するように直接的に訓練されるかどうかについては、ほとんど知られていなかった3。私たちや他の研究者は、微生物やワクチンへの呼吸器粘膜の曝露が、TII14,15あるいは免疫制御性や寛容性を有する気道メモリーマクロファージを誘導することを発見しています16,17,18。呼吸器アデノウイルスワクチン/感染症は、肺胞マクロファージ(AM)に、循環単球とは無関係に持続的なメモリー表現型を誘導した。このようなメモリーマクロファージに関連するTIIは、肺の標的および異種細菌病原体の両方に対する自然保護機能を強化する14,19.
したがって、免疫学的刷り込みやトレーニングの歴史に由来する安静時の自然免疫記憶の形成において、コンパートメント化のパラダイムを支持する証拠が増えてきているのである。このパラダイムでは、訓練された造血前駆細胞や循環単球は全身的/両親からの微生物曝露/ワクチン接種に起因し、一方、バリア粘膜常在マクロファージの記憶表現型は局所微生物曝露/ワクチン接種に起因するとしている3,20,21。後者は、マクロファージの組織内滞留ニッチとその局所炎症への適応という現在の概念に合致するものである22。局所的な炎症がない場合、全身的/非全身的な微生物曝露/ワクチン接種後に遠距離から発生する免疫学的シグナルを統合し適応する方法として、バリア組織での自然記憶が発達するかどうかは不明なままであった23。最近の研究では、皮下ウイルス感染/ワクチン接種が、複数の組織部位にまたがる広範な免疫学的警告を引き起こすこと24,25、局所組織損傷が遠隔組織部位の常在マクロファージを活性化すること26が示されている。組織部位間の免疫学的クロストークの一つの典型的な様式は、腸内細菌叢の異常が肺の免疫応答を変化させる腸肺軸である27, 28, 29, 30, 31, 32, 33, 34. しかし、非経口ワクチン接種が腸内細菌叢に影響を与えるかどうか、腸内細菌叢の異常が肺の自然記憶を誘導するかどうか、またどのように誘導するかはよく理解されていない23。BCGは、現在のほとんどのヒトワクチン(コロナウイルス病2019(COVID-19)ワクチンを含む)と同様に皮膚から投与されるため、これらの疑問に取り組むことは、広範囲な意味を持つ。
ここでは実験モデルを用いて、BCG皮下接種が肺の組織常在メモリマクロファージとTIIを誘導するかどうか、またどのように誘導するかを検討した。BCGは骨髄前駆細胞に対する効果とは別に、結核菌感染に対するメモリーマクロファージとTIIを非経口的に誘導することが知られている。この過程は、最初のマイコバクテリアの拡散と、腸内細菌叢、バリア機能、代謝産物の時間依存的変化を伴う腸-肺軸を介して起こる。このように、我々の研究は、腸内細菌叢を介した遠位粘膜部位での自然免疫記憶の発達経路を明らかにし、呼吸器系病原体に対する次世代ワクチンの開発に示唆を与えるものである35,36。
結果
BCG皮下接種により肺マクロファージに記憶表現型が誘導される
BCG皮下接種により、BM/血液中の訓練された単球が誘導される9,13。しかし、肺に存在するAMが訓練されるかは不明である。そこで、マウスにBCGを皮下接種した。ワクチン接種後5週目に、気管支肺胞洗浄液(BAL)および肺単核細胞を、結核菌全細胞溶解液(WCL)による刺激あり(S)または刺激なし(US)で解析した(図1a)。AMはLy6G-CD11b-CD11chiCD64hiSiglec-Fhi骨髄系細胞として同定された14。BCGを接種した宿主の気道細胞の約95%がAMであった。コントロールのAM(PBS-US)と比較して、BCG AM(BCG-US)は刺激により高レベルのベースラインおよび増強されたMHC IIを発現した(PBS-S vs BCG-S)(図1b)。また、刺激されたAMは、有意に増加したtoll-like receptor 2 (TLR2)を示した(図1c)。刺激されたBCG AMのIL-6産生頻度は有意に高く、腫瘍壊死因子(TNF)産生細胞は同等であった(図1d,e)。対照宿主とBCG接種宿主の肺組織AM集団の間で、同様の免疫プロファイルが観察された(拡張データ図1a-c)。
図1:皮下BCGワクチンにより遠位呼吸器粘膜にメモリーAMが誘導される。
図1
b,c, 気道常在菌のMHC II (**P = 0.0027, ****P < 0.0001) (b) またはTLR2 (**P = 0.0050) (c) の表面発現のヒストグラム. d,e, IL-6- (***P = 0. 0007) (d) またはTLR2のヒストグラムと頻度. 0007)(d)またはTNF産生気道AM(e)。 f、(S)または(US)刺激なしで培養した気道AMの上清中のサイトカイン/ケモカインのタンパク質レベル(幾何平均値)のヒートマップ。赤いアスタリスクは、BCG宿主の気道AMによる刺激でサイトカイン/ケモカイン産生が有意に増加したことを示す。 g-l、IL-6 (*P = 0.0230) (g), IL-12p40 (****P < 0.0001) (h), TNF (i) and MCP-1 (**P = 0. 0031) (j) の濃度は、BCG宿主の気道AMによる刺激で、IL-6、IL-12p40、TNF、MCP-1の濃度は、0. 0031) (j), MIP-1α (*P = 0.0408) (k) and RANTES (*P = 0.0137) (l) in airway AM cultured with and without stimulation. m, Real-time ECAR in airway AM at 8 weeks post-BCG immunization. 2-DG: 2-deoxy-glucose. n, BCG免疫後8週間の気道AMにおける解糖(***P = 0.0006), 解糖能 (***P = 0.0007) および解糖予備能 (**P = 0.0013). o、BCG接種後5週目のPMによるF4/80表面マーカーを発現する全生体腹膜細胞のうちゲートされた細胞の頻度およびF4/80発現の蛍光強度中央値(MFI)のドットプロット(*P = 0. 0231、***P = <0001)。 p、MHC IIのMFIと、生BCGまたは不活化BCG(BCG-ia)またはPBSで(S)または(US)刺激を加えたワクチン接種後5週間におけるIL-6またはTNF産生PMの頻度のヒストグラム(***P = <0.0001)。b、dおよびeのデータは、3つの独立した実験の代表値である。f、g-lおよびo-pのデータは、2つの独立した実験の代表である。棒グラフのデータは、平均値±s.e.m.として示され、生物学的に独立したサンプルの個々のデータポイントを表し、n = 3マウス/グループである。b-lのデータにはボンフェローニ検定を用いた一元配置分散分析、nのデータには両側t検定、oとpのデータには一元配置分散分析とダネットの多重比較検定が用いられた。
フルサイズ画像
分泌されたサイトカイン/ケモカインの11種類のLuminex分析では、ベースライン(US)では、サイトカイン/ケモカインのレベルはコントロールおよびBCG AM培養物で低かった(図1f-l)のに対し、刺激すると、コントロールAMと比較して、BCG AMは有意により高いレベルのIL-6, IL-12p40, MCP-1, MIP-1α およびRANTESを生成し(図1g、h, j-l) TNF生成では差が観察できなかった(図1i).
次に、気道AMの代謝状態について解析した。BCG AMは、酸化的リン酸化速度が軽度に変化したのに比べ、解糖が著しく増加した(図1m、nおよびExtended Data図1d)。局所的なウイルス感染で鍛えられたAMは、維持のために低速のin situ増殖を行うため14、BCG AMの増殖能をin vivo BrdU取り込みで調べた(Extended Data 図1e)。BCG AMはコントロールAMに比べて有意にBrdU取り込みを増加させ(Extended Data Fig.1f)、BrdU+およびBrdU-AMの両方がMHC IIの増加を示したため、その増加は増殖状態とは無関係だった(Extended Data Fig.1g )。
次に、BCG後2週間の早い時点で、気道AMの免疫表現型と代謝状態を調べた(Extended Data Fig.1h)。5週目に獲得した訓練された表現型とは対照的に、BCG宿主の気道および肺組織のAMの表現型は対照群と同等であった(拡張データ図1i-k)。また、2週間後のBCG宿主では、対照群に比べ解糖が有意に低下していた(Extended Data Fig.1l)。このような時間依存的な気道AMの訓練のメカニズムを評価するために、まず、BCG後のマイコバクテリアの播種との関係を検討した。2週間後、縦隔リンパ節(MLN)へのBCG播種がわずかに認められたが(40 ± 20 colony forming unit (CFU) / MLN)、肺への播種は認められなかった。BCGは倍加時間が遅いので、この所見は記憶型AMの誘導に時間依存的なマイコバクテリアの播種が関与していることを示唆するものであった。次に、BCGの生存・複製と播種がAMの訓練に果たす役割を、生存BCGと熱不活性化BCG(BCG-ia)の皮下注射で比較検討した。BCG接種後の生存宿主のAMとは異なり、BCG-ia AMは未訓練のAMと同様にMHC IIとIL-6の発現プロファイルを示した(Extended Data 図1m)。
また、BCG接種が肺以外に腹膜腔のマクロファージにグローバルな影響を与えるかどうかを調べた。腹膜マクロファージ(PM)は、CD11b+F4/80hiSiglecF-集団として同定された37。BCG PMは、PBSコントロールと比較してF4/80発現が減少し(CD11b+F4/80LowSiglecF-)、再刺激なしで(US)構成的に増加したMHC II発現とex vivo再刺激で(S)IL-6/TNF生産が増加する鍛えられた表現型(図1o)と著しく変化した免疫表現型を示している(図1p)。逆に、BCG-iaはAMを訓練できなかったのと同様に(拡張データ図1m)、PMを訓練することもできなかった(図1p)。
以上のデータから、非経口BCGは時間依存的に記憶型AMの誘導を引き起こすことがわかった。このようなメモリーAMは、刺激によりMHC IIやTLR2の発現、解糖、サイトカイン産生が増加することが特徴である。また、BCG接種により腹腔内のマクロファージがグローバルに養成される。
メモリー肺マクロファージによる特異的な遺伝子プロファイルと微生物制御
次に、BCG宿主のメモリーAMの転写プロファイルを調べた。気道BALのBCGまたはコントロール細胞を、(S)刺激あり、または(US)刺激なしで培養し、単離したRNAを配列決定した(図2a)。主成分分析(PCA)により、各グループは独自のパターンにクラスタリングされ、それぞれに固有の遺伝子発現プロファイルがあることが示された(図2bおよびExtended Data 図2a)。ベースラインとWCL再刺激後のBCGとコントロールAM集団の間で、合計248個の遺伝子が差次的に発現(DE)していた(Extended Data 図2b,c)。BCG AMは、刺激の有無にかかわらず、細胞周期と分裂に関与する遺伝子セットに富んでいた(図2cおよび拡張データ図2d)。例えば、細胞増殖マーカーであるMki67(Ki67)、染色体分離や紡錘体形成に関わるキネシン様タンパク質をコードするKif11、Kif15、Kif23、DNA修復に関わるRad51、Rad54b、姉妹染色体分離に関わる遺伝子はBCG AMでアップレギュレーションされていた。また、BCG AMは防御応答遺伝子、特にT細胞の走化性(Cxcl10、Ccl5)、単球(Ccl7)に関与する遺伝子をアップレギュレートし、細胞活性化遺伝子Lck、Bcl2、IL7r、Slamf7をダウンレギュレートした(Fig. 2d)。さらに、TII1および抗原(Ag)の処理と提示14、解糖、mTOR経路および脂肪酸酸化に関連する定義済みの遺伝子セット(補足表1)を、PBS(US)対BCG(US)、PBS(S)対BCG(S)で比較した。MHC IIの増加(図1b)に伴い、抗原提示に関わる遺伝子の発現量は、ベースライン(US)および刺激時(S)にBCG AMで有意に増加したが(図2e)、解糖およびmTOR経路のものは有意差がなかった(拡張データ図2e,f)。一方、脂肪酸の酸化に関連する遺伝子の発現は、刺激したBCG AMで有意に減少し(図2fおよび補足表1)、酸化から解糖へと代謝がシフトしていることと一致した(図1m,n)12。これらのデータは、BCGワクチン接種がメモリーAMのユニークな転写プロファイルを引き起こすことを示唆している。
図2 BCGワクチンによるメモリーAMの転写と抗菌反応。
図2
a, 実験スキーマ b, 気道AMの遺伝子発現のPCA, n = 3マウス/群 c, PBSとBCG宿主の気道AMの細胞周期に関与するDEGのヒートマップ。e,f, 気道AMのAg処理および提示(*P = 0.0260; **P = 0.0054)(e) および脂肪酸酸化(**P = 0.0076)(f) に関与する遺伝子のシグネチャースコア(Signature score)。g, 気道AMによる結核菌のex vivo貪食/殺傷の実験スキーマ。 h, 24時間(**P = 0.0005)および48時間(**P = 0.0006)で評価した気道AMによる貪食された結核菌の殺傷率。各ポイントは生物学的に独立したサンプルを表す(PBS群ではn = 4マウス、BCG群ではn = 3マウス)。i、BCG-dsRed感染後4時間(貪食)および24時間(殺傷)におけるdsRed+気道AMの頻度の代表的フローサイトメトリープロット。 j、BCG-dsRed感染後4時間および24時間(**P = 0.0072) における気道AM内のdsRedシグナルのMFI。各ポイントは生物学的に独立したサンプルを表す(各群n = 3マウス)。 l, 結核感染前の気道AMにおけるMHC II発現のヒストグラムとMFI(***P = 0.0010)。各ポイントは生物学的に独立したサンプルを表す(1グループあたりn = 3マウス)。調整後のP値は、バイオリンプロットで示され、limmaパッケージとBH補正(方法)を使用して得られた(e,f)。統計分析は、h、j、およびlのデータについて、BCG(US)とPBS(US)およびBCG(S)とPBS(S)を比較する両側t検定により決定された。
フルサイズ画像
次に、BCG AM対PBS AMによる差次的発現遺伝子(DEG)に関する本研究のデータを用い、経鼻的にアデノウイルス(Ad)ワクチンを接種したマウスのAMにおけるDEGと比較して解析した14。その結果、BCG AM(Extended Data Fig. 2b)と比較して、訓練されたAd AM(Extended Data Fig. 2g)では約5倍の遺伝子(1,309対248)が差次的に発現していることが判明した。しかし、抗原提示、解糖、mTOR経路、脂肪酸酸化に関連する事前定義された遺伝子セットでは、両者とも同様の特徴を有していた(Extended Data Fig.2h)。Ad AMとBCG AMを区別するトランスクリプトームの特徴は、細胞活性化遺伝子、Lck、Bcl2、Il7r、Slamf7がアップレギュレートされ、細胞周期および分裂関連遺伝子が濃縮されないことであった。これらのデータは、訓練されたAMのいくつかの特徴は、特定のワクチン戦略間で共有されているが、ワクチンの種類と送達経路によって訓練されたAMはユニークであることを示唆している。
BCGで訓練されたAMは、MHC IIおよび抗原提示/処理遺伝子が増強されていることから、Ag85Bを添加したBCG AMと共培養したトランスジェニック結核菌Ag85B特異的CD4 T細胞を使って、T細胞への抗原提示能力をex vivoで評価した(Extended Data 図3a)。T細胞の増殖率は、FACSによるCFSE希釈の程度として校正された。BCG AMと培養したT細胞の約30%が少なくとも3世代増殖したのに対し(G)、対照AMと培養したT細胞の15%はほとんど1世代増殖にとどまり(拡張データ図3b,c)、BCG AMによって抗原提示が促進されていることが示された。
抗菌活性を調べるために、BCGおよびコントロールの気道AMを結核菌にex vivo感染させ、CFUアッセイによりマイコバクテリアの阻止/殺傷率を測定した(Fig. 2g)。BCG AMは、対照AMと比較して、感染後24時間および48時間の両方で結核菌の制御能力(図2h)、または蛍光タンパク質を発現するMycobacterium bovis(BCG-dsRed)の制御能力(図2i・j)が有意に優れていることが示された。BCG肺組織のCD11C+/CD11b+抗原提示細胞(APC)でも結核菌制御の増強が見られた(拡張データ図3d)。訓練されたAMとコントロールのAMは、どちらも同様の貪食率(図2i、j)および細胞死/アポトーシス率(拡張データ図3e)を示した。MHC IIを介した生体外での結核抗原提示の増加(Extended Data 図3b、c)と同様に、訓練を受けたAMは、生体内結核菌感染によりMHC II発現を急速に増加させた(図2k、l)。これらのデータは、BCG宿主における記憶表現型に加えて、訓練されたAMは、独特の転写プロファイルを示し、抗原提示と抗マイコバクテリア活性を増加させることを示している。
肺結核に対する記憶肺マクロファージによる訓練された免疫力
抗結核宿主防御は、長い間、非経口的BCGワクチン接種によって誘導される適応的Th1免疫のみに起因すると考えられてきた。気道AMは結核菌の大部分を保有し、結核の初期段階での播種に寄与することが知られている38。BCGで訓練された気道AMによる生体外環境での菌制御/反応の増強(図2h-l)は、このようなAMが生体内のT細胞免疫とは別に結核に対するTIIを提供できることを示唆している。記憶型AMが抗結核TIIを提供するかどうかを検討するために、4週間のBCGワクチン接種を受けた動物を結核に感染させ、7日目と14日目に肺CFUを評価した(Fig.3a)。ワクチン未接種対照と比較して、BCG肺では7日目と14日目にそれぞれ約0.3 logと約1 logの結核菌CFUが減少していた(Fig. 3b)。気道AMが最初の7-9 d38内の主要なM.tuberculosisリザーバーであることと一致して、3日目と7日目のBCG気道細胞(BAL細胞)(図3c)は、非接種対照と比較してM.tuberculosis CFUが著しく減少し(図3d)、3日目のBCGホストの無細胞液(BALF)(図3e)中のM. tuberculosis CFUも著しく減少することと一致する。これらのデータは、BCGで訓練されたAMがin vivoでよりよく結核を制御できることを示唆している。
図3:メモリーAMは、感染初期に肺結核に対する訓練された免疫を付与する。
図3
a, 実験スキーマ b, 結核感染後7日および14日の肺組織CFU (dpi) (*P = 0.0342; **P = 0.0047).7dpi(PBS群n=8マウス;BCG群n=9マウス)および14dpi(PBS群n=4マウス;BCG群n=5マウス)の2つの独立した実験からプールされた生物学的に独立したサンプル。c、実験スキーマ。d,e、3dpi(n=5マウス/群;*P=0.0208)および7dpi(n=4マウス/PBS群;n=5マウス/BCG群;*P=0.0476)における気道内の細胞内(BAL細胞)結核菌CFU。 0476)(d)、および3dpi(PBS群n=5マウス;BCG群n=4マウス;*P=0.0152)および7dpi(各群n=5マウス)における気道の細胞外(BALF)結核菌(e)。 f、実験スキーマ。 g、3、7および14 dpiにおける活性化マーカー CD44および Ag85B テトラマーに陽性な気道 CD4 T細胞の代表点プロット。h,i, 3 (n = 3マウス/グループ)、7 (n = 4マウス/グループ)、14 (n = 4マウス/グループ) dpiにおける気道のCD4+CD44+ (*P = 0.0490; **P = 0.0016) (h) およびCD4+tet+ T細胞 (***P = 0.0003) (i) の合計数. j, 0, 3 (*P = 0.0162), 7 (**P = 0.0082) および 14 (*P = 0.0260) dpi における LPT の CD4+CD44+ T 細胞の総数. k,l, 代表的なドットプロット(k)および感染後のLPTにおけるCD4+tet+ T細胞の総数(***P = 0.0002)(l). m, 実験スキーマ n, 7 dpiにおける肺結核CFU (*P = 0.0438; **P = 0.0013; ****P ≤ 0.0001).p,q, 3dpiにおける気道(*P = 0.0404) (p) と肺組織(*P = 0.0308) (q) の細胞内 (BAL 細胞) M. Tuberculosis CFU 数、n = 4 マウス/グループ/組織。散布図における横線は、s.e.m.エラーバー付きの平均を示す。統計解析は、BCG群とPBS群を比較するb、d、e、h-j、l、pおよびqについては両側t検定により決定し、nのデータは一元配置分散分析、続いてフィッシャーの最小有意差(LSD)検定により解析した。
フルサイズ画像
次に、BCG宿主における結核菌のAM制御の強化におけるBCG活性化Th1-cellの潜在的役割について調査した。まず、Ag85B-CD4 T-cell tetramers (tet) 39 を用いて、M. tuberculosis 後早期の時点における気道および肺実質組織 (LPT) の抗原特異的 CD4 T-cell の動態を明らかにした (Fig. 3f)。LPT内のT細胞は、血管内CD45.2免疫標識40を介して血管内対応物と区別された(Extended Data Fig. 4a)。実質的に活性化されたCD4+CD44+ T細胞は、感染後3日目という早い時期にBCG気道に現れ、一方、コントロール気道には7日目まで現れなかった(図3g,h)。同様に、tet+CD4 T細胞(CD4+Ag85B+)は7日目にBCG気道にのみ見られたが、対照気道には見られなかった(図3g,i)。しかしながら、CD4+CD44+およびCD4+Ag85B+ T-細胞の両方は、BCGおよびコントロール宿主の間で14日目に同等となった(図3g-i)。同様に、BCG LPTは、対照と比較して3、7および14日目に有意に大きな活性化CD4+CD44+ T-細胞の数を有し(図3jおよび拡張データ図4a)、また特に7日目により多くのCD4+Ag85B+ T-細胞を含んだ(図3k、l)。これらのデータは、訓練されたAMによる結核制御の強化が、結核感染の初期に気道内のAg特異的T細胞の増加を伴っていることを示唆している。
訓練されたAMと結核菌制御の増強との直接的な関係を明らかにするために、結核菌感染前のBCG宿主のT細胞をmAbを用いて枯渇させ(図3m)、BCG肺中の7日目の結核菌CFU (BCG ΔT細胞)とワクチン未接種(PBS)および対照Ab処理したBCG宿主(BCG)のそれを比較検討した。先のデータ(図3b)と一致して、BCG肺(BCG)には有意に減少したM.tuberculosis CFUが含まれていた(図3n)。BCG宿主のT細胞(BCG ΔT細胞)の枯渇は、増強された防御力を損なうことなく、むしろM. tuberculosis CFUをさらに減少させた(Fig. 3n)。ワクチン未接種(PBS)宿主のT細胞の枯渇は、M. tuberculosis CFUに影響を与えなかった(PBSの4.27±0.06に対してPBS ΔT細胞では4.16±0.08)。逆に、養子移入されたBCG AM(BCG-AM)を受けたナイーブ動物は、対照AM(PBS-AM;拡張データ図4b)を受けた動物と比較して、肺のM. Tuberculosis CFUが中程度に減少していた。次に、古典的なLy6Chi単球を欠くCCR2KO結核感染モデルを用いて、BCG宿主41における採用単球由来AM(MDM)の役割を検討した(図3o)。BCGワクチンを接種したCCR2KOマウスは、ワクチンを接種していないコントロールに比べて、気道マクロファージ(図3p)および肺組織(図3q)の両方で結核菌に対して有意に良好な防御能を維持した。以上のデータから、BCG宿主における結核の初期防御の増強は、T細胞や循環単球とは無関係であり、訓練されたAMの役割をさらに裏付けていることがわかる。
メモリーマクロファージ誘導における単球とT細胞の独立性
BCG非経口接種では、骨髄前駆細胞の刷り込みによって循環単球が訓練されることが示されている9,13ので、我々のモデルで骨髄系細胞と循環単球が訓練されているかどうかを調べた(図4a)。実際、BCG宿主では、MPP3骨髄系前駆細胞の頻度がコントロールに比べて有意に増加していることが観察された(図4b)。ゲーティング戦略(拡張データ図5a)を用いると、BCG宿主の末梢血中のLy6ChiおよびLy6Clow単球はいずれも、刺激により対照と比較して高レベルのMHC IIを発現することがわかった(図4c、dおよび拡張データ図5b)。ただし、IL-6の産生能には違いがなかった(拡張データ図5c)。一方、BCG宿主のLy6Clow単球は、刺激によりTNFを産生するようになり(図4e)、TNF+Ly6Chi単球は同等であった(Extended Data 図5d)。循環単球の活性化の増強と一致して、BCG宿主からの刺激された全血培養物におけるIL-1β、IL-6、IL-12p40、IP-10、MIP-1αおよびRANTES産生は対照と比較して著しく増加した(図4fおよびExtended Data 図5e)。これらのデータは、BCG皮下接種がBMにおける骨髄造血の増加をもたらし、循環単球を訓練することを示す。
図4:メモリーAMsの誘導は、訓練された循環単球およびT細胞由来のシグナルに依存しない。
図4
b, 骨髄系(MMP3)およびリンパ系(MMP4)前駆細胞の代表的ドットプロット、およびBMの全多能性前駆細胞のうちMMP3の頻度(*P = 0.0120)(n = 3マウス、n = 4マウス、PBS群) c,d, 循環Ly6Chigh(*P = 0.0228) (c) およびLy6Clow(P = 0. 0510) (d) のMHC IIのMFI。 e、Ly6Clow TNF+単球の頻度(S)、(US)刺激なし(*P = 0.0157)(n = 3マウス/群)。 f、全血培養サンプルの血漿中サイトカイン/ケモカイン蛋白レベル(幾何平均)のヒートマップ(S)、(US)刺激なし。g, h, SiglecF+Ly6C- airway AM (g) と肺組織 (h) の代表的ドットプロットである。肺組織中のMDMとIMはそれぞれSiglecF-Ly6C+とSiglecF-Ly6C-として同定された。気道および肺組織におけるマクロファージサブセットの総数を棒グラフに示す。2つの独立した実験の代表(n = 3マウス/グループ/組織)。i、BCGワクチン接種またはPBS処理したCCR2KOマウスの気道AM上のMHC IIのMFIは刺激あり(S)および刺激なし(US)、気道AMの培養上清中のサイトカイン/ケモカインレベルは刺激あり(S)(*P = 0. j, BCG接種またはPBS処理したCCR2KOマウスの肺組織AMにおけるMHC IIのMFI(US刺激あり(S)、US刺激なし(S):*P = 0. 0238; S: *P = 0.0246)、刺激の有無による肺組織AMのIL-6産生頻度(US: *P = 0.0284; S: *P = 0.0297) およびTNF産生頻度を示す。l, BCGワクチン接種またはPBS処理したWTおよびCCR2KO動物の気道におけるPKH標識AMの代表的ヒストグラム、PKH標識のない(PKHなし)ナイーブマウスAMと比較。 m,n, PBS および BCGワクチン接種宿主の気道AMにおける胚由来(AM)(m)と循環単球遺伝子(n)のシグニチャースコア。バイオリンプロットの横線は中央値を、点線は下位および上位四分位を示す。 p, 刺激のあるAMの培養上清におけるサイトカイン/ケモカインのタンパク質レベル(幾何平均)のヒートマップ。赤色のアスタリスクは、BCG宿主の気道AMを刺激した際の有意差を示す。 q,r, 気道AM上のMHC IIのMFI (**P = 0.0066; ****P ≤ 0.0001) (q) およびIL-6産生気道AMの頻度 (*P = 0. 0159; ****P = 0. 0159). 0159; ***P = 0.0003; ****P ≤ 0.0001) (r) を、PBS、BCG-ワクチン接種群およびBCG/IFN-γ欠失(抗IFNγ)群を比較して、(S)刺激ありおよび(US)刺激なしと比較した。棒グラフのデータは、平均±s.e.m.として示される。統計分析は、BCGとPBSを比較するb-e、i、およびjについて、両側t検定によって決定された。qおよびrのデータは、一元配置分散分析、続いてボンフェローニ検定による多重比較によって分析した。
フルサイズ画像
循環単球は、特に肺の炎症状態において、AMのプールに寄与している可能性がある21,22。BCG宿主におけるメモリーAMの発生と訓練された循環単球の関係に取り組むため、まず、気道の炎症性サイトカイン/ケモカインのレベルを評価したところ、すべて検出されなかった。次に、気道のマクロファージと単球の表面マーカーを調べた。BCG後2週間(拡張データ図6a)および5週間(図4g)において、BCGおよびコントロールの両ホストのほとんどの気道(BAL)マクロファージはSiglec-F+ resident AMであった。さらに、主要なマクロファージ集団である単球由来マクロファージ (MDM) (Siglec-F-Ly6C+) 、間質性マクロファージ (IM) (Siglec-F-Ly6C-) および AM (Siglec-F+Ly6C-) の頻度と総数は、両群の肺組織でほぼ同じだった (Extended Data 図 6b および 4h)。したがって、これらのデータは、BCG宿主における気道常在記憶型AMの誘導に対する循環単球の実質的な寄与を支持しない。
これをさらに調べるために、古典的なLy6Chi単球を欠いたCCR2KOマウスを5週間BCGワクチン接種した。コントロールと比較して、BCG接種CCR2KO動物の気道および肺組織のAMでは、刺激の有無にかかわらずMHC IIの発現が上昇したままであった(図4i,j)。さらに、刺激すると、コントロールと比較して、CCR2KO宿主からのBCG AMは、有意に高いレベルのIL-6およびTNFを産生した(図4i)。野生型(WT)BCG AMのサイトカインプロフィールと同様に(図1d,e)、刺激したCCR2KO BCG AMは、TNF産生細胞は同等であったが、IL-6を産生する頻度が有意に高かった(図4j)。BCG CCR2KO動物由来のAMの訓練された表現型/免疫性は、これらの細胞が結核菌のex vivo感染により、有意に良好に結核菌感染を制御したという機能データによってさらに裏付けられた(拡張データ図6c)。これは、ex vivo感染WT BCG AM(図2g-j)およびBCG CCR2KO動物の肺の抗結核TII(図3o-q)とも一致していた。これらの知見は、BCGワクチン接種によって、循環単球に依存しない機能的メモリーAMが誘導されることを示すものである。
別のアプローチで、安定な蛍光色素PKH26をWTおよびCCR2KOマウスの両方に投与し、貪食42を介して気道常在AMを標識した。BCG訓練されたAMに対する循環単球の寄与があれば、気道AM内のPKH26は希釈されたであろう(図4-k)。ワクチン非接種のWT(WT-PBS)およびCCR2KO(CCR2KO-PBS)ホストでは、気道AMの大部分は5週間にわたってPKHによって安定的に標識されたままであった(図4l)。WTおよびCCR2KO BCG宿主のAMからのPKHの損失/希釈はなく(図4l)、BCG訓練されたAMに対する循環単球の寄与は最小であることが示唆された。自律的に誘導されたAMは、その胚性(AM)および単球性遺伝子シグネチャー14において定常状態のものと類似した状態を保つであろうから、これらの遺伝子を対照とBCGで比較したところ、実質的な違いは見られなかった(図4m、n)。したがって、記憶AM誘導における循環単球の独立性がさらに支持されることになる。
BCGは単球/マクロファージの訓練に関与するIFN-γを産生するTh1細胞を活性化する(図3g、I、k、l)ため、BCG訓練されたAMにおけるT細胞とIFN-γの役割を明らかにした。T細胞やIFN-γは、BCG後2週間の訓練されたAMの欠如時からmAbを用いて枯渇させ(Extended Data 図1h-l)、その枯渇はその後3週間にわたって維持された(図4o)。以前の観察(図1f)と一致するが、BCG AMは刺激により多量のサイトカインを産生し(S)、T細胞枯渇は再刺激に対するBCG AMによるそのような増強された応答を損なわず、むしろIL-1β、IL-6、MIP-1αおよびMIG産生のさらなる増加をもたらした(図4p)。同様に、IFN-γ中和は、MHC II(図4q)、IL-6(図4r)およびTNF(拡張データ図6d)産生の上昇を伴う訓練されたAM表現型を損なわないことが判明した。これらの結果は、BCGワクチン接種された宿主におけるT細胞枯渇が、結核の初期段階における防御の増加を損なわなかったという事実と一致する(図3n)。以上のデータから、BCGによる気道常在記憶型AMの誘導/維持は、訓練された循環単球、T細胞ヘルプまたはIFN-γに依存しないことが示された。
腸内細菌叢、代謝産物、バリア機能の変化
最近のエビデンスによると、腸内細菌叢は遠位感染に反応して変化し、そのような変化は腸-肺軸を介して肺の免疫応答を変化させる可能性がある27,28,29,30,31,32,34。我々のモデルにおいて、腸-肺軸が粘膜常在型メモリーAMの誘導に関与しているかどうかを調べるため、まず5週間後の盲腸のマイクロバイオームを特徴付けた。BCG接種により、5週目の盲腸の大きさは有意に減少した(Extended Data Fig.7a)。操作的分類単位(OTU)に基づくα多様性比較では、BCG盲腸において微生物の豊かさまたはα多様性指数が有意に低いことが明らかになった(図5a)。細菌門のランクにおける実際の存在量もBCG動物で減少し、上位4つの門の平均存在量は対照動物の33,125に対して20,329(P = 0.06)だった(拡張データFig.7b)。BCG宿主の腸内細菌叢は、主座標分析(PCoA)では一部重複していたものの、対照動物とは有意に分離していた(P < 0.004, PERMANOVA; 図5b)。さらに、細菌科のランクでは、S24-7としても知られるMuribaculaceaeが対照動物とBCG宿主の両方で優勢だったのに対し、BCG宿主の腸内マイクロバイオームではLactobacillaceaeの頻度が著しく高かった(図5c,d)。BCG宿主では、19個の特定のOTUが差次的に変化していた(偽発見率(FDR)<0.05;補足表2)。AMにおけるTIIの有意な誘導は5週目に観察されたが(図1a-p)、2週目には観察されなかったので(拡張データ図1h-l)、BCG後2週目の腸内マイクロバイオームの特徴も調べた。実際、2週間の腸内細菌叢はあまり変化せず、コントロールの腸内細菌叢に似ていたが(拡張データ図7c-f)、部分的に重複し(拡張データ図7e)、14の特定のOTUが差次的に変化した(補足表3)。これらの結果は、BCG接種後の腸内細菌の時間依存的な発達を示唆している。
図5 BCG接種後の腸内細菌叢および微生物代謝産物の変化。
図5
a,b, 盲腸内細菌叢におけるChao1多様性指標を用いたOTU豊富度に基づく5週間PBS群とBCG接種群のα多様性比較(t検定)(P = 0.0540) (a) およびPCoA順序法およびJenson-Shannon発散距離法(PERMANOVA)によるβ多様性比較(P < 0.0040) (b). n = 6マウスPBS群, n = 5マウスBCG群. c,d, 上位8ファミリーの相対的存在量プロファイルの円グラフ(c)およびCMにおける上位4ファミリーの頻度の比較棒グラフ(P = 0.0505)(d). n = PBS群6マウス、n = BCG群5マウス. e,f, H&E染色結腸切片の代表顕微鏡写真(n = PBS群6マウス、n = BCG群5マウス)。g、BCG後の指定された時点における大腸の建築的変化、上皮の変化、および炎症性浸潤の組織学的スコアリング(n = 5/2, 6/5, および 4/8 週)。i、経口投与したFITC-デキストランの循環動態の光学濃度として測定した腸管透過性の比較(*P = 0.0012)。j, 5週間のPBSおよびBCG宿主の糞便組織サンプルにおける代謝プロファイルのPLS-DA分析。k, 5週間のPBSおよびBCG宿主の糞便組織サンプルにおけるデオキシカルニチンレベル(湿重量1グラム当たりの相対ピーク面積)の比較(*P = 0. l, 5 週間の PBS と BCG 宿主の血清サンプルにおける代謝プロファイルの PLS-DA 解析。m, 5週齢のPBSおよびBCG宿主の糞便組織サンプル中のSCFAsの総濃度(mM g-1)のうち、酪酸SCFAの割合を算出した(P = 0.0512)n = 16マウス/群. n, PBS、2週齢および5週齢BCG宿主の肺組織サンプル中の代謝プロファイルに対するPLS-DA分析の比較. o,p, カルニチン生成物の比較:ブチルカルニチン(***P = 0.0007)()とヘキシルカルニチン()。 PBS、2-/5-週BCG宿主の肺組織サンプルにおけるブチリルカルニチン(*P = 0.0007)(o) およびヘキサノイルカルニチン(*P = 0.0268)(p; 湿重量1グラムあたりの相対ピーク面積)。 r, 2週間BCG宿主のMLN、腹膜細胞画分(PCL)中の無細胞PW中のBCG CFU数. a-dのデータは、2つの独立した実験の代表値である。j-mのデータは、2つのプールされた実験から得られたものである(1グループあたりn = 16マウス)。箱ひげ図における横線は中央値を、箱の長さは下位および上位四分位を、ひげは最小値および最大値を表す。棒グラフのデータは平均値±s.e.m.で示され、円グラフの数値は中央値頻度を表す。統計解析は、d、i、k、mおよびqのデータについて、BCGとPBSを比較する両側t検定によって決定された。oとpのデータは、一元配置分散分析、次いでボンフェローニ検定による多重比較によって分析した。
フルサイズ画像
次に、BCG宿主における腸内細菌異常が、ワクチン接種後5週間における顕微鏡的組織学的変化を伴うかどうかを評価した。コントロールと比較して、BCG宿主の遠位結腸では、不規則な絨毛、短縮した陰窩、拡大した内腔、上皮の破壊/剥離、粘膜の炎症性浸潤を特徴とする著しい変化が認められた(図5e、fおよび拡張データ図7g)。これらの変化は明瞭であるが、ネズミの腸の炎症に関するガイドラインによれば、重症度は軽度であった43。BCGワクチン接種によって誘発された大腸炎が時間依存的で自己限定的であるかどうかを調べるために、BCG後2週間および8週間における大腸組織学を調べ、大腸炎が顕在化した5週間と比較した。2週間後の腸内細菌叢の変化は小さく(Extended Data Fig.7c-f)、大腸組織学は低度のリンパ球浸潤を除いて変化がなかった(Fig.5gおよびExtended Data Fig.7h)。5 週間後の大腸炎(Fig.5e,f)とは対照的に、接種後 8 週間までに大腸炎はほとんど消失し、大腸の構造も回復した(Fig.5g and Extended Data Fig.7h).これらの知見は、非経口BCGが時間依存的ではあるが自己限定的な腸炎を誘発することを示している。5週間後の大腸炎を想定して、上皮のタイトジャンクション蛋白、zonula occludens (ZO-1) および occludin の発現を免疫組織化学的に検討した。BCG宿主の大腸上皮では、ZO-1(図5h)とオクルーディン(拡張データ図7i)の分布が、対照宿主では均一/無傷であるのに対し、不規則で乱れた状態であることが確認された。その結果、フルオレセインイソチオシアネート(FITC)標識デキストラン法(図5i)で示されるBCG宿主の腸管透過性が有意に上昇していることが確認された。これらの知見は、腸内構造変化と腸管内腔分子の上皮横断移動の増加との関連性を示唆するものである。
腸内細菌症はしばしばその代謝物の変化と関連していることから28 、5 週間の BCG 動物から得た盲腸、結腸および血清のメタボロームをプロファイリングした。Partial least square-discriminant analysis (PLS-DA)の結果、盲腸の代謝物はグループ間でクラスタリングされ、一部重複していることがわかった(Fig. 5j)。対照群に比べ、BCG宿主の盲腸では、L-カルニチンの前駆体であるデオキシカルニチン/γ-ブチロベタイン(図5k)およびクレアチニン(拡張データ図8a)の濃度が有意に上昇した。また、各群の大腸の代謝物もそれぞれのパターンに分かれており(Extended Data 図8b)、BCG宿主では乳酸レベルが大幅に上昇していた(Extended Data 図8c)。腸内細菌代謝物の変化の他に、PLS-DAモデルは血清代謝物の明確な群間クラスタリングを示した(図5l)。VIPスコア(>1.5)に基づき、10種類の血清代謝物を順位付けしたところ、クレアチンを除くすべての代謝物の濃度が、BCG宿主では対照群と比較して低下していることが分かりました(Extended Data 図8d、右側のカラーボックス)。マウス血清メタボロームのパスウェイ解析により、アルギニン代謝経路が対照群と比較してBCG宿主で主に影響を受けていることが明らかになった(Extended Data 図8e)。短鎖脂肪酸(SCFA)は、食物繊維の分解に伴って生成される腸内細菌叢の主要代謝物のひとつであり、腸内外で炎症/代謝に影響を及ぼすことから28、主要SCFAを定量化した。BCGによる腸内環境の異常によって盲腸の酢酸は影響を受けなかったが、プロピオン酸と酪酸の相対的な割合は変化し(Extended Data 図8f)、酪酸レベルは有意に増加した(図5m)。PICRUSt44を予測ツールとして用い、BCGと対照動物との違いに基づく腸内細菌叢の機能的可能性についても検討した。エネルギー代謝、トランスポーター、シグナル伝達と細胞プロセス、遺伝情報処理に分類される19の予測機能分子が有意に異なった(P < 0.05)(補足表4)。盲腸内の酪酸レベルの上昇(図5m)に伴い、ブトン酸(酪酸の塩およびエステル)代謝に関与するアセト乳酸合成酵素I/II/IIIラージサブユニットがBCG宿主で有意に増加した。代謝物以外にも、微生物関連分子パターン(MAMPs)を含む他の管腔内分子も、透過性の増加により循環系に移行する可能性があるため、血清LPSレベルを測定したが、両者に違いは見られなかった(<0.01 EU per ml; Extended Data Fig.8g)。
BCG宿主の腸/血清におけるメタボロームの変化を考慮し、2週間および5週間後の肺のメタボロームを調査した。腸内細菌/メタボロームの2週間後(拡張データ図7c-f)および5週間後(図5a-d,j-m)の変化がそれぞれほとんどないことと同様に、肺の代謝物のプロファイルは5週間後でのみ有意に異なり、2週間後では見られなかった(図5n)。VIPスコア(>1.5)に基づき、7つの肺代謝産物がランク付けされた(Extended Data 図8h)。5週目の盲腸におけるデオキシカルニチンレベルの上昇と同様に(図5k)、カルニチン生成物であるブチリルカルニチンおよびヘキサノイルカルニチンはBCG後5週目に肺で有意に上昇した(図5o,p)。これらのデータは、腸、血清、肺の間のメタボローム変化における時間依存的な関連性を示している。
肺のAM以外にも、BCG接種はPMに対してグローバルなトレーニング効果があり(図1o,p)、腹膜腔・マクロファージと腹膜腔内の腸を含む臓器との間に生物学的関係が存在することを考え、BCG接種が腸の変化をどのように媒介するかを調べるために、BCG後2週間の腹膜腔へのBCGトランスロケーションの可能性を検討した。実際、結腸の長さは対照とBCG宿主で有意差があり、後者は〜2 cm短縮しており、軽度の結腸炎を示唆していた(図5q)だけでなく、かなりのBCG CFUがMLN、無細胞腹膜洗浄(PW)および全マクロファージ画分(腹膜細胞溶解液(PCL))においてのみ検出され(図5r)、未接種対照からは検出されなかった。これらのデータは、遠位型BCGワクチン接種と腸の顕著な変化との間の機構的連関を立証するものである。
以上のデータから、BCG接種は、マイコバクテリアの腸管関連部位への播種を介して、腸内細菌叢、メタボロームおよびバリア機能の時間依存的変化をもたらし、その結果、血清および肺のメタボローム変化ももたらすことが示唆された。
BCGワクチンで調整した腸内細菌叢が肺マクロファージを記憶させる
次に、BCGによる腸内細菌叢の異常とメモリーAMの誘導の関係について、微生物叢移植のアプローチを用いて検討した。ナイーブマウスを広域抗生物質で10日間処理して元の微生物叢を撹乱してから、対照(PBS-CM)またはBCG(BCG-CM)宿主からの腸内細菌叢(CM)を移植し、その後5週間にわたって定着させた(図6a)。BCG AMの訓練された表現型(図1a-n)と同様に、BCG-CMでコロニー形成された気道AMは、PBS-CMでコロニー形成されたものと比較して、ベースライン(US)および刺激時(S)にMHC II、IL-6、TNF産生の上昇を実証していた(図6b-d)。同様の訓練された表現型は、PBS-CMではなく、BCG-CMの肺組織でAMでも見られた(図6e-g)。したがって、これらのデータは、BCGで調整した微生物叢と記憶型AMの誘導との間の因果関係を立証するものである。
図6:BCGワクチンで調整した腸内細菌叢の移植により、メモリーAMが誘導され、免疫力が鍛えられる。
図6
a, 実験スキーマ b, c, d, BCG-CM群からの気道AMによる訓練された自然免疫特性。気道AMのMHC II (*P = 0.0477) のMFI (b), IL-6 (**P = 0.0046; **P = 0.0025) (c) およびTNF (*P = 0.0109, *P = 0.0192) (d) の生成頻度、刺激あり (S) および刺激なし (US) の気道AM。n = PBS-CM群5匹、n = BCG-CM群4匹。e,f,g, BCG-CM群の肺組織AMによる訓練された自然免疫の特徴。肺AMのMHC II (**P = 0.0052) のMFI (e), IL-6 (P = *0.0122) (f) およびTNF (*P = 0.0144) (g), 刺激あり (S) およびなし (US) の肺AMの生成頻度. h、BCG-CMマウスの結腸における上皮過形成、杯細胞の減少および軽度の上皮崩壊を示すH&E染色結腸切片の代表顕微鏡写真(PBS-CM群のn = 5マウス、BCG-CM群のn = 4マウス)。i、MUC2タンパク質を免疫組織化学的に染色した結腸切片の代表的な顕微鏡写真(PBS-CM群n=5マウス、BCG-CM群n=4マウス)。赤いアスタリスクは、PBS-CMマウスと比較して、BCG-CMマウスの結腸上皮におけるMUC2染色の有意な減少を示す。 j、PBS-CMマウスからのものと比較して、刺激を与えたBCG-CMマウスからの循環Ly6Chi (****P < 0.0001) およびLy6Clow (***P = 0.0002, **P = 0.0011) 単球上のMHC IIのMFI。l,m,n, 細胞内(BAL細胞)(*P = 0.0431) (l) および細胞外(BALF) (m) のM. tuberculosis CFUの数(PBS-CM群n=5、BCG-CM群n=6)および肺組織中のCFU数(n)(PBS-CM群n=4、BCG-CM群n=6)は、感染後3日目にPBS-CMマウスと比べ、BCG-CMマウスでは、気道のCFUは、PBS-CM群で5、PBS-CM群で6、BCG-CM群で6であった。統計解析は、PBS-CM(US)対BCG-CM(US)およびPBS-CM(S)対BCG-CM(S)を比較したb-g、jおよびlのすべてのデータについて、両側t検定によって決定された。
フルサイズ画像
予想通り、BCGワクチン接種宿主(図5e、fおよび拡張データ図7g)とは異なり、ナイーブ動物のBCG-CMコロニー形成は、大きな結腸構造変化を引き起こさなかった(図6h)。しかし、PBS-CM対照と比較して、杯細胞の減少と軽度の上皮崩壊を伴う上皮過形成(図6h)および上皮ムチン-2(muc-2)産生の減少をもたらした(図6i)。腸内細菌症はBM骨髄造血の変化と関連しているので47、循環単球を調べると、BCG-CMコロニー化がPBS-CMコントロールに比べて循環Ly6ChighおよびLy6Clow単球を有意に活性化することがわかった(図6j)。
BCG-CMコロニー形成で鍛えられたAMが、BCGワクチン接種した宿主に見られるような肺TIIに移行するかどうかを調べるために(図3b、d、e、n、p、q)、CMコロニー形成したナイーブ動物を結核菌に感染させて3日目にCFUを評価した(図6k)。実際、気道BCG-CM AM(BAL)は、PBS-CM対照と比較して有意に減少したマイコバクテリアを含んでいた(Fig.6l)。これに対応して、無細胞BALFおよび肺組織中のCFUも、BCG-CM動物において小さい傾向にあった(図6m,n)。以上のデータを総合すると、BCGで調整した腸内細菌叢を単独で移植すると、腸管バリアの変化、メモリーAM、肺の抗結核TIIが誘導されることが示された。
肺マクロファージの訓練における循環微生物代謝産物の役割
BCGで訓練されたAMと循環代謝産物の関係を調べるために、単球の訓練用に確立されたin vitroモデルを適応した48。ナイーブな気道AMを、BCGワクチン接種動物または対照動物の血清を添加した培養液で培養した(トレーニング)。24時間のトレーニング後、24時間または3日間の静止後、細胞を刺激し、顕微鏡で分析し、免疫型を決定した(図7a)。コントロール血清とは異なり、BCG調整血清(BCG-S)は、トレーニングまたは休息後、および再刺激時にAMの著しい形態変化を引き起こした(図7bおよびExtended Data図9a)。これらのAMはクラスターに集まり、細胞質包有物を伴って大きくなった(訓練後;図7b)。安静時には、細胞分裂と拡散が増加したように見えた(安静後)。これは、BCG宿主のAMで細胞分裂に関与する遺伝子が濃縮されていることと一致している(図2cおよび拡張データ図2b-d)。最も顕著な形態学的変化は、再刺激時(刺激後)に見られ、有意に上昇したMHC IIおよびIL-6産生を伴っていた(図7c-e)。この訓練された免疫表現型は、BCGワクチン接種した宿主の記憶AMと類似しており(図1b,d,e)、3日間休ませたAMでのみ観察され、訓練後24時間休ませたものでは観察されず、以前の観察と一致した48。さらに、トレーニングバイオマーカーのIL-1β、IL-6、TNF、およびMCP-1とKCの産生は、BCG-Sに曝露したAMから、コントロールに比べて有意に増加した(図7f、g)。これらのデータは、BCG訓練を受けたAMにおける循環可溶性因子の役割を示唆している。
図7:BCG接種を受けた宿主の循環微生物代謝産物は、AMの自然免疫トレーニングにつながる。
図7
a, AMのin vitro自然免疫トレーニングの実験スキーマ。 b, BCG-Sによるトレーニングまたはコントロール血清による処理後、あるいは再刺激後のAMの代表的な明視野顕微鏡画像。c-e、BCG-Sで訓練したAMと24時間または3日間の休息後の再刺激で、MHC II (*P = 0.0132 および *P = 0.0182) (c) およびIL-6 (*P = 0.0362) (d) の頻度(TNF (e) では蛍光強度(MFI)中央値が上昇した。各データポイントは、PBS血清あたりn = 3ウェル、BCG血清あたりn = 4ウェルを表す。 f,g, BCG-Sで訓練したAMの培養上清および3日間静置後の再刺激時のサイトカイン/ケモカイン蛋白質含有量を示す。各データポイントは、各条件につきn = 4ウェルを表す。TNF, **P = 0.0047; KC, *P = 0.0491. h-j, メチル基転移酵素阻害剤MTAまたはアセチル基転移酵素阻害剤EGCGでヒストンのメチル化およびアセチル化を阻害した際のBCG-SによるAMの自然免疫訓練の抑制. データは、再刺激時にAMによって産生されるIL-1β (***P = 0.0008) (h), IL-6 (***P = 0.001) (i) およびTNF (****P < 0.0001) (j) のタンパク質含有量を示している。各データポイントは、各条件につきn = 4ウェルを表す。統計解析は、c-eのデータについては、刺激後の各時点におけるPBS血清(US)対BCG血清(US)およびPBS血清(S)対BCG血清(S)の比較、fおよびgのデータについては、PBS血清(S)対BCG血清(S)、h-jについては、BCG-Sとの比較で、t検定の結果両側で決定されている。
フルサイズ画像
自然界のトレーニングには、ヒストンのメチル化/アセチル化を介したエピジェネティックな再プログラミングが関わっていることから1、AMに対するBCG-Sのトレーニング効果の観察に、ヒストンの修飾が関与しているかどうかを評価した。そこで、BCG-Sによるトレーニングの間、培養液にヒストンメチルトランスフェラーゼ阻害剤5′-deoxy-5′-methylthioadenosine(MTA)またはヒストンアセチルトランスフェラーゼ阻害剤epigallocatechin-3gallate(EGCG)48を補充した。ヒストン修飾酵素、特にヒストンアセチルトランスフェラーゼの阻害(BCG + EGCG-S)は、再刺激時の BCG-S 訓練 AM による IL-1β, IL-6 および TNF 産出を有意に減少させた(図 7h-j)。メチルトランスフェラーゼ阻害剤(BCG+MTA-S)では、比較的小さな抑制効果が観察された。これらのデータは、BCG宿主の循環代謝産物によるAM訓練におけるエピジェネティック修飾の関与を示唆するものである。
腸内細菌由来の循環系SCFAs28の免疫調整的役割と、BCG接種宿主で観察されたデオキシカルニチンおよび酪酸の増加(図5j-p)を考慮し、これらのSCFAsと記憶型AM誘導の関係を明らかにした。L-カルニチンと酪酸塩酸塩の混合物をナイーブ動物の飲料水(DW + M)に3週間導入し49 、対照動物には通常の飲料水(DW)を与えた(Fig. 8a)。DW + MのAMは、刺激(S)により、調べたサイトカインのうちIL-6とMIP-1αの産生が対照(DW)よりも増加し(図8b、cおよび拡張データ図9b)、BCG訓練したAMのプロファイル(図1g、k)と同様であることが示された。しかし、BCG宿主の循環単球(図4fおよび拡張データ図5)とは異なり、DW + M単球は訓練された表現型をとらず、Ly6CおよびMHC II発現が低下して抑制された免疫プロファイルを示した(拡張データ図9c,d)。サイトカイン/ケモカインの分泌能力も同程度であった(Extended Data Fig.) さらに、予想通り、BCG接種宿主とは異なり(Fig. 5e,f および Extended Data Fig. 7g)、代謝物補給(DW + M)は軽度のリンパ球浸潤を除いて大腸建築の変化を引き起こさなかった(Extended Data Fig.9f)。
図8:BCGワクチンを接種した宿主の微生物代謝産物を補給した水は、ナイーブマウスのAMの自然免疫トレーニングにつながる。
図8
a、ナイーブマウスに3週間与えた(DW+M)を介して循環SCFAs酪酸およびL-カルニチンのin vivo連続補給の実験シェーマ。マウスはその後、通常のDWに1週間置いた。b,c、DW + MまたはDWで、(S)または(US)再刺激を行った動物から単離した気道AMによるIL-6(*P = 0.0071)(b) およびMIP-1α(*P = 0.0381)(c) のタンパク質産生。e, 対照(DW)と代謝物添加(DW + M)動物の気道AMにおけるAg処理および提示に関与する遺伝子のシグネチャスコアを、(US)刺激あり(S)となし(P = 4.34 × 10-6)で比較。バイオリンプロットの横線は中央値を、点線は下位および上位四分位を示す。 f、代謝物補給水(DW+M)を与えられたナイーブ動物の肺組織における結核菌CFUの数とBCGワクチン接種(DW+M vs BCG、*P = 0.0203)(BCG vs PBS *P = 0. 0115)の比較を行ったマウス。 g-j, WT (**P = 0078 and *P = 0.0118) (g), TLR2KO (**P = 0. 0043) (h), TLR2KOの気道AMにおけるMHC II発現のMFIを比較した結果, PBS群では3マウス、BCG群では4マウス、DW + M群では4マウスが、DW = 0.0115でした。 0043)(h)、TLR4KO(i)およびNOD2-KO(**P = 0.0062)(j)マウスをBCG調整またはコントロール血清でex vivo訓練し、3日間休息後の再刺激時に、気道AMのHHC II発現を測定した。棒グラフのデータは平均値±s.e.m.で示した。統計解析はb、c、fのデータについて両側t検定により行った。調整後のP値はバイオリンプロット(e)について示し、limmaパッケージとBH補正を用いて求めた(Methods)。統計解析は、g-jのデータについて、PBS血清(US)対BCG血清(US)およびPBS血清(S)対BCG血清(S)を比較する両側t検定により決定された。
フルサイズ画像
次に、メタボロームサプリメントとBCGワクチン接種によって訓練されたAMのトランスクリプトームプロファイルを比較した。DW + M気道AM(図8a)を、刺激あり(S)および刺激なし(US)で転写解析した。各グループは独自のパターンに集積しており、代謝物処理に伴う AM の転写変化が示唆された(Fig. 8d)。DW + M AMでは、DW対照と比較して、合計265個の遺伝子が差次的に発現していた(Extended Data 図9g、h)。BCG AM(図2cおよび拡張データ図2d)と同様に、細胞の分化/増殖に関連する遺伝子(Nov、Hbegf、KitlおよびSix5)も、DW + M AMでは対照と比較して発現が上昇した(拡張データ図9g)。炎症反応に必要な微小器官/ミクログリア活性化遺伝子であるSncaも、両方のAMで発現が上昇した(図2dおよびExtended Data 図9g)。また、BCG AM(図1bおよび図2c,e)と同様に、HLA遺伝子(拡張データ図9h)および抗原提示遺伝子(図8e)を含む免疫遺伝子は、刺激したDW + M AMで有意に増加した。さらに、DW + M AM の脂肪酸酸化、解糖および mTOR 経路に関連する遺伝子(拡張データ図 9i-k) のレベルは、概ね BCG AM のレベル(図 2f および拡張データ図 2e、f)と同様であった。重要なのは、DW + M肺では、結核菌感染の初期段階において、BCG宿主よりもさらに大きな範囲で有意に増強された保護があったことである(図8f)。これらの知見は、BCG変化代謝物の補充のみで、BCGワクチン接種と同様にAMの訓練された免疫表現型、トランスクリプトーム変化およびTIIを誘導し得ることを示すものである。
血清LPSが検出されなかったことから(拡張データ図8g)、BCGワクチン接種後の宿主において循環系に移行する可能性のある腸内MAMPsの役割についてさらに検討した。微生物のペプチドグリカン、LPS、ムラミルジペプチドがそれぞれTLR2、TLR4、NOD2のリガンドであることを考慮し、TLR2-、TLR4-、NOD2-欠損(KO)AMに、当社のex vivo AMトレーニングモデルを使ってBCG-SまたはPBS調整済み血清でex vivoトレーニングを行いました(図7a)。BCG-Sは、WT AMの無刺激(US)および有刺激(S)において、自然界における訓練マーカーであるMHC IIの増加を有意に誘導したが(図8g)、無刺激のTLR2-またはNOD2-KO AMにおいてもMHC IIの増加を誘導した(図8h,j)。M. tuberculosis WCLで刺激すると、NOD2-KO AM、特にBCG-Sで訓練したAMで、MHC IIがさらに増加した(図8j)。興味深いことに、TLR4-KO AMは、刺激前のコントロールとBCG血清の間で差のない、低下したMHC IIを発現していた(図8i)。刺激によってこれらの細胞でMHC IIが増加したが、それはコントロールとBCG-Sの間で差がなかったことから、AMによるMHC II発現にTLR4シグナル伝達が本質的に必要であることが示唆された。上記のデータは、たとえ循環レベルの上昇した状態で存在しても、これらのMAMPは大きく寄与しないことを示唆している。しかし、循環微生物代謝産物は、BCGワクチン接種によるAMトレーニングにおいて重要な役割を担っている。
考察
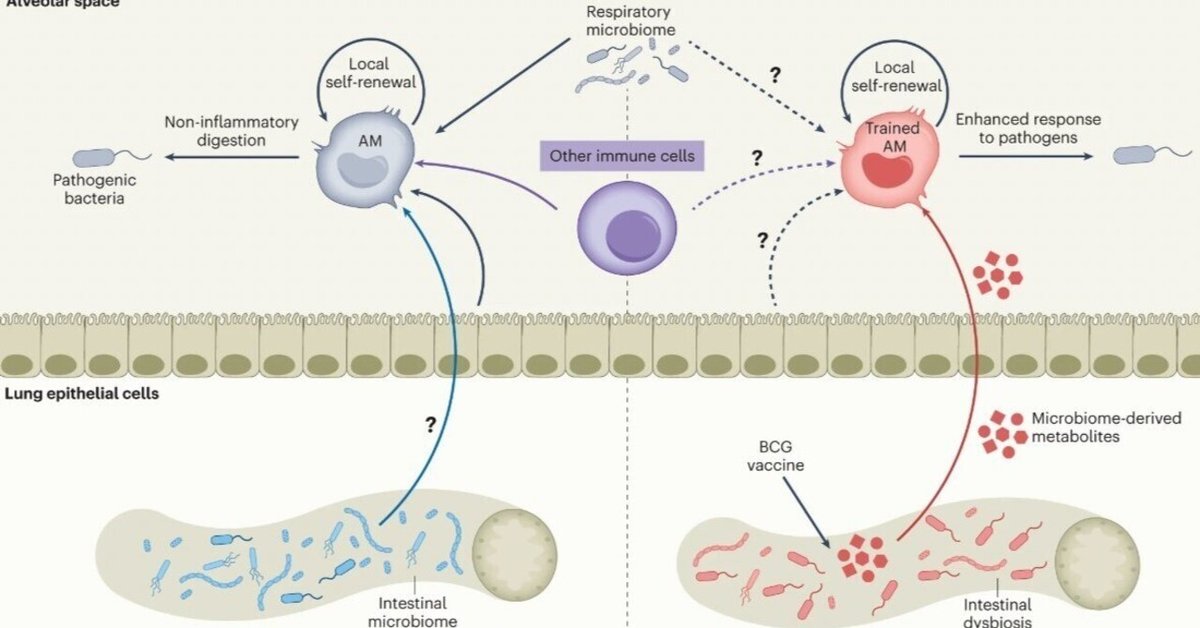
ワクチン接種後、局所炎症がない場合、遠方で発生した免疫学的シグナルを統合し適応することにより、肺に存在する自然免疫記憶が生じるかどうかは依然として不明である。本論文では,BCG皮下接種が,循環単球に対する訓練効果に加えて,時間依存的にメモリーAMおよびTIIを誘導することを示した.このようなメモリーAMは、循環単球とは無関係に発達し、自己維持される。BCG接種は、マイコバクテリアの移動、腸内細菌の異常増殖、透過性の増加、局所/全身代謝物の変化などを含む腸-肺軸を介してこれを行う(Extended Data 図10)。
このように、我々の研究は、非経口ワクチン接種後の遠位粘膜部位における自然免疫記憶/TII発達のための新しい腸内細菌代謝経路を明らかにし(拡張データ図10)、自然免疫記憶の形成は免疫学的曝露経路によって区分されるという現在の見解を変えるものである2,3,20。このように、非経口ワクチン接種は、複数の組織部位にまたがる長距離の免疫学的警告を引き起こし、結果としてマクロファージメモリーを形成する可能性がある。このような知見は、非経口ワクチンによる宿主防御機構に対する我々の理解を深めることになる。適切に設計された非経口ワクチンの接種後、病原体が呼吸器に侵入すると、訓練された粘膜常在マクロファージが宿主防御の第一線として働き、訓練された循環単球の動員によって強化されるという、「正規組織訓練免疫」と呼ばれる機構が示された20。また、BCG接種を受けたヒトの肺では、非特異的自然保護が強化されるという新たなメカニズムも提供されている5,6,7,9. BCG非経口接種と異なり、アデノウイルス非経口接種ではAM19を訓練できないことは注目に値する。これは、粘膜常在TII誘導にはワクチンプラットフォームと送達経路の選択が重要であることを示唆している。BCG-iaがAMを訓練できないことから、非経口ワクチンの複製性が、広範囲の免疫学的警告とグローバルなマクロファージ訓練効果に必要であることが示唆され、このことは、腸-肺軸を開始するためにBCGの伝播が必要らしいという我々の発見によってさらに裏付けられた。BCGはゆっくりと複製されることから、BCGがゆっくりと広がり、腸内細菌の異常、大腸炎、メタボロームシフトが時間依存的に発現することが説明できるかもしれない。重要なことは、軽度の大腸炎は8週間までにほぼ消失するため、自己限定的であるということである。最近、BCG非経口接種に伴う腸内細菌叢の変化を示す2つの報告があるが50,51、非経口BCG関連大腸炎に関する臨床報告はなく、腸内のメタボロームシフトや肺のTII誘導との関連は言うに及ばずである。BCG接種に伴う大腸炎は軽症で一過性であるため、臨床的な意義がないものと思われる。さらに、非経口 BCG 接種を受けたヒトのうち、肺で結核菌に対する自然保護機能を獲得するのはごく一部であることから52、腸-肺軸を介した肺残留型抗結核 TII の誘導は、おそらく遺伝的に決定されていて、一部のヒト BCG ワクチン接種者にのみ発生するものである。
また、非経口ワクチン接種により、腸内細菌叢に関連した局所的・全身的なメタボローム変化が引き起こされる可能性があることも明らかにした。さらに、腸-肺軸を介した肺内在性自然免疫記憶の誘導は、T細胞やIFN-γとは無関係であり、局所炎症経路を介した肺粘膜内在性マクロファージの形成における中心的役割とは異なることを証明した14。肺への影響に加え、腸内細菌の代謝異常は、我々の現在の研究で示されたように、おそらく骨髄造血に影響を与えることで循環単球を鍛える可能性もある47。腸内細菌叢由来のSCFAは免疫調節特性を有しており23,33、インフルエンザ発症前のマウスでは、優勢なSCFAである酢酸の減少がAMの殺菌性に影響を与え30、SCFA産生の欠損がミクログリアの成熟を阻害していた49。腸肺のデオキシカルニチンおよび酪酸レベルの上昇がAMトレーニングに関連しているという我々の発見は、SCFAsの自然制御特性に関する我々の理解を大きく深めるものである。肺のTIIを誘導するためにSCFAsを飲料水から補給するという我々のアプローチは、免疫治療戦略の可能性を示している。今後、非経口BCGをきっかけとした腸内環境の異常が、肺や腹腔以外の粘膜組織でマクロファージを訓練する可能性があるかどうかを検討する必要がある。
また、我々の発見は、肺のホメオスタシスと宿主防御における組織常在型AMの可塑性を強調するものである3,21。AMの訓練性、耐久性、自律性は、這うように肺胞を巡回し、好中球よりも高い割合で細菌を死滅させる能力と一致する53。このように、我々は、BCGワクチン接種を受けた宿主において、T細胞や循環単球とは無関係に、メモリーAMが結核の防御力を高めることを示した。これは、BCG接種を受けたヒトのかなりの割合で観察される、結核菌の自然除去の基礎となるもっともらしいメカニズムである52。結核菌は気道マクロファージを乗っ取って播種する能力があることから、BCG接種者の結核菌侵入部位に訓練されたAMがいることは、早期感染制御のために重要である19,38,39。結核菌以外にも、非経口 BCG による肺 TII が異種細菌感染に対する防御になるかどうかを現在調査しているところである。
結論として、我々の研究は、非経口ワクチンをきっかけとした腸内細菌叢を介した遠位粘膜組織における自然免疫記憶発達への新たな経路を明らかにした。また、生ワクチンの非経口免疫により、中枢性と末梢性の両方でTIIが誘導されることが示された。このような知見は、結核やSARS-CoV-2などの呼吸器系病原体に対する次世代ワクチンの設計に役立つと考えられる(参考文献35,36)。
研究方法
マウス
WT雌のC57BL/6マウスは、Charles River Laboratories (Saint Constant) またはJackson Laboratory (Bar Harbor)から購入した。雌のケモカイン(C-Cモチーフ)受容体2(CCR2)(B6.129S4-Ccr2tm1Ifc/J)、TLR2(B6.129-Tlr2tm1Kir/J)、TLR4(B6(Cg)-Tlr4tm1.2Karp/J)、NOD2(B6.129S4-Tlr4tm2Kir/J)。129S1-Nod2tm1Flv/J)ノックアウトおよびC57BL/6バックグラウンドでAg85B受容体(H2-Kb-Tcra,-Tcrb)P25Ktk/J)を発現するCD4 T細胞を含むP25 TCR-Tg transgenic mouseはJackson Laboratoryから購入した。すべてのマウスは到着時に6-8週齢であった。マウスは、マクマスター大学の特定病原体不検出レベルB施設またはバイオセーフティレベル3施設において、餌と水に自由にアクセスでき、12時間の光サイクル、50〜60%の湿度、20〜25℃の室温で飼育された。各実験では、同じ部屋に収容された年齢を一致させたマウスを使用した。コントロールマウスには、BCGの調製に使用したPBSを皮下投与した。動物は無作為に実験群を割り当てた。すべての実験は、McMaster UniversityのAnimal Research and Ethics Boardsからの機関指針(AUP 210822)に従って実施された。
BCGによる皮下免疫
マウスを4〜5×104CFUのBCGパスツール株54で皮下免疫した。熱BCG-iaは、4-5×104CFUのBCG投与量を85℃の水浴で45分間インキュベートすることによって調製した。不活性化は、コロニー形成アッセイで確認した。
M. tuberculosisによる肺感染
強毒性(H37Rv;ATCC 27,294)および無毒性(H37Ra;ATCC 25,177)のM. tuberculosisの感染量を既報19と同様に調製した。マウスにM. tuberculosis H37Raを0.5 × 106 CFU/マウス、またはM. tuberculosis H37Rvを1 × 104 CFU/マウスで感染させた。M. tuberculosis H37Rvの負荷は、肺ホモジネート、BAL液、または滅菌水で破壊して細胞内マイコバクテリアを放出したBAL細胞の連続希釈物を3連でプレーティングして、指定したエンドポイントにおいて評価された。
気管支肺胞洗浄および腹膜洗浄と単核細胞の単離
マウスは失血死させた。場合によっては、失血の3分前に抗CD45.2抗体(クローン104)(BD Pharmingen)40をi.v.注射して血管内染色を実施した。BAL および肺組織中の細胞は、既報14,40 に従って分離した。腹膜腔は、以前に記載されたように洗浄した37。簡単に言えば、3mlの全洗浄液(2mM EDTA、1mM HEPESを含むPBS)を腹膜腔に注入し、腹膜を30秒間穏やかにマッサージし、体壁に小さな切り込みを入れてピペットチップで腔液を採取した。脾臓単核細胞は、以前に記載された方法で入手した40。BM細胞は、脊椎、大腿骨および脛骨をPBS中で乳鉢で粉砕することにより得た。その後、BM細胞を40μmのバスケットフィルター(BD Biosciences社製)で濾過した。赤血球を溶解した後、分離した細胞をex vivo培養用に完全RPMI 1640培地(10%FBSおよび1%L-グルタミンを添加したRPMI 1640、1%ペニシリン/ストレプトマイシン添加または無添加)またはフローサイトメーター染色用にPBSに再懸濁させた。細胞内サイトカイン染色(ICS)のためにBALおよび肺細胞を刺激した場合、または培養上清中のサイトカイン/ケモカインレベルを測定するために培養した場合、細胞は2%FBSを含む完全RPMI 1640培地に再懸濁させた。
免疫染色、in situ 細胞増殖およびフローサイトメトリー
細胞の免疫染色とフローサイトメトリーは、以前に記述したように実施した14。具体的には、肺胞マクロファージ活性化レベルおよび細胞内サイトカイン産生を測定するために、BALからの単核細胞25万個および肺組織からの単核細胞2×106個を平底48ウェルプレートにプレートし、マクロファージの付着および新鮮に分離したAMの無関係な炎症性活性を和らげるために37℃で3時間インキュベートした。インキュベーションの終わりに、非接着細胞を洗い落とし、1.6μg ml-1の濃度のM.tuberculosis WCLを含む、または含まない新鮮な培地を加えた。訓練された循環単球のレベルおよび細胞内サイトカイン生産を測定するために、心臓穿刺によりEDTA血液チューブ(サルステッド)に全血を集め、等量のRPMI 1640で希釈した。希釈した全血を300μlに分注し、1.6μg ml-1の濃度でM. tuberculosis WCLとともに、またはM. tuberculosis WCLなしにインキュベートした。GolgiPlug (5 mg ml-1) (BD Biosciences) を、刺激剤を添加した1時間後に、BALおよび肺細胞、ならびに希釈全血培養物に添加した。PMの活性化レベルを決定するために、1×106個の単核細胞を、1.6μg ml-1の濃度のWCLと共に、およびWCLなしでU底96ウェルプレートにプレーティングした(参考文献55)。刺激剤を添加した1時間後にGolgiPlugを添加し、さらに5時間培養した。刺激終了後、冷蔵庫で氷冷したFACS buffer(0.5% bovine serum albumin)(Sigma Aldrich)中で15分間インキュベートしてBALおよび肺細胞をウェルから浮揚させた。EDTA(Sigma-Aldrich)(30 μl of 20-mM EDTA per tube)と共に室温で15分間インキュベートし、BD Pharm Lyse(BD Biosciences)を用いて赤血球を溶解後、免疫染色用の全血単核球を得た。Aqua dead cell staining kit (ThermoFisher Scientific) で染色後、細胞を洗浄し、抗CD16/CD32 (clone 2.4G2) でブロックし、BD Cytofix/Cytoperm (BD Biosciences) で製造者の指示に従って固定および透過処理を行った。場合によっては、BALおよび肺単核細胞を培養せずに免疫染色し、表面メーカーの発現とイムノフェノタイピングを行った。
マクロファージおよび単球におけるTIIの特徴付けに使用した蛍光色素標識mAbは、補足表5に記載されている。骨髄系(MMP3)およびリンパ系(MMP4)前駆細胞13に偏光する多能性前駆細胞の同定には、mAbのパネルを使用した(補足表5)。T細胞のICSのために、BAL、肺、脾臓細胞をGolgiPlug(5 mg ml-1 brefeldin A; BD Pharmingen)存在下で、粗BCGと結核菌培養ろ液の混合物(2 μg per well)54を添加または無添加で培養した。刺激した細胞を細胞表面抗体で染色し、固定/透過化溶液キット(BD Biosciences社製)を用いて、製造者の指示に従って固定/透過化した。その後、Perm/Wash buffer(BD Biosciences)中の抗IFN-γ-APC mAbで氷上で30分間染色した。T細胞表面およびICSに使用した蛍光色素標識mAbは、補足表5に示した。テトラマー免疫染色には、C57/Bl6 MHCクラスIIアレルに結合したAg85Bの免疫優位CD4 T細胞ペプチド(FQDAYNAAGHNAVF)のテトラマー(PE蛍光色素に結合したI-A(b)(Ag85B:H-2I-AB)(NIH Tetramer Core, Atlanta, GA)39が使用された。
in situ AM増殖の測定には、APC BrdU flow kit (552598; BD Biosciences)を用いた。BrdUの経鼻投与は、BCG免疫後5週目に繰り返し行い、合計9日間、マウス1匹あたり0.5mg、総量50μlの濃度で投与した14。その後、DNAに取り込まれたBrdUを抗BrdU-APC mAb (clone B44)で検出した56。
特に断りのない限り、すべてのmAbと試薬はBD Biosciences社から購入した。すべての抗体は、実験に使用する前に、最適な条件を確認し滴定した。免疫染色された細胞は、BD Biosciencesのフローサイトメトリーの説明書に従って処理し、FACSDivaソフトウェアを使用してBD LSR IIまたはBD LSRFortessaフローサイトメーターで実行された。データはFlowJoソフトウェア(バージョン10.8.1; Tree Star)を用いて解析した。
ケモカイン・サイトカイン定量
BAL AMが産生するサイトカインおよびケモカインを測定するために、2%FBS含有完全RPMIに懸濁した細胞を96ウェル平底プレートに10万個/ウェルでプレーティングし3時間静置した。細胞を洗浄後、1.6μg ml-1のM. tuberculosis lysateを添加または無添加した2%FBS含有完全RPMIをウェルに加えて37℃、5%CO2で12~14時間インキュベートした。回収した培養上清は、タンパク質量を測定するまで-80℃で凍結した。全血を等量のRPMIで希釈し、400μlを48ウェル平底プレートにプレートし、1.6μg ml-1の結核菌溶解液で刺激するかしないかで、37℃、5% CO2で12-14時間インキュベートした。その後、血漿を採取し、タンパク質レベルの測定まで-80℃で保存した。サイトカイン/ケモカインレベルは、MCYTOMAG-70Kマウスケモカインおよびサイトカイン検出キット(Millipore Sigma, Oakville, ON)を用いて、製造者の指示に従って定量化した。プレートをMagPixリーダーで読み取り、サイトカイン/ケモカインの濃度をxPONENTソフトウェア(Thermo Fisher Scientific)を使用して決定した。
Ex vivoでのマイコバクテリアの貪食および殺傷アッセイ
マイクロビーズ(Miltenyi Biotec)を用いて、BALのAMと肺から精製したプールしたCD11b+細胞およびCD11c+細胞のマイコバクテリア貪食および殺傷アッセイは、前述19と同様に実施した。細菌殺傷能力(% killing)を計算し、式により培養プレート内のCFUを測定した。殺滅率=(4時間後の細菌CFU数-24/48時間後の細菌CFU数)/(4時間後の細菌CFU数)×100.さらに,AMのマイコバクテリア貪食・殺傷能力についても,赤色蛍光を発現する組み換えBCG(BCG-dsRed)およびフローサイトメトリーにより評価した。マイコバクテリウム感染細胞(dsRed+)のアポトーシスおよびネクローシスは、Aqua dead cell staining kitおよびAnnexin V-APC staining kit(BD Biosciences)で細胞をメーカー指示通りに染色後、フローサイトメーターで解析した。
Ex vivoでの肺胞マクロファージによるCD4 T細胞への抗原提示
P25-Tg マウスの脾臓およびリンパ節から精製した Ag85B 特異的トランスジェニック CD4 T 細胞を Carboxyfluorescein succinimidyl ester (CFSE) (Invitrogen) で標識した39。CFSE標識したCD4 T細胞をBALから得たAMと2:1の割合(2×105 T細胞対1×105 AM)で37℃、5%CO2で96時間共培養し、細胞を洗浄、CD3およびCD4細胞表面マーカーを免疫染色し、CD3+CD4+細胞のCFSE希釈度をフローサイトメトリーで分析した。
In vitroマクロファージ養成とエピジェネティックな再モデリング抑制
ナイーブマウスのBALから得たAMを%L-グルタミンを含むRPMI1640に懸濁し、1%ペニシリン/ストレプトマイシンを添加または無添加で、プレート(96ウェルでは10万個/ウェル、48ウェルでは25万個/ウェル)にして37℃、5%CO2で3時間安静とした。自然免疫訓練は、5週齢のBCG免疫マウスまたはプラセボマウスから得た血清を各ウェルに最終容量の2%添加して24時間培養した。訓練後、細胞を培養液(%L-グルタミン含有RPMI1640、1%ペニシリン/ストレプトマイシンおよび2%FBS含有または含有しない)で一度洗浄してから24時間または3日間培養液中で静止させた。休息後、M. tuberculosis WCL(1.6 μg ml-1)で12-14時間刺激した。培養上清中のサイトカイン/ケモカインタンパク質をLuminexで測定した。別に、ウェルから上げた細胞を免疫染色し、フローサイトメトリーで解析した。エピジェネティック修飾酵素の阻害が自然免疫訓練に影響するかどうかを調べるために、BCG血清中で24時間インキュベートした細胞に、1mMの濃度のヒストンメチルトランスフェラーゼの阻害剤、5′-deoxy-5′-MTA(Sigma-Aldrich)または15μMの濃度のヒストンアセチルトランスフェラーゼ、EGCG(Sigma-Aldrich)を添加した48.
In vivoでのT細胞の枯渇とIFN-γの中和
In vivoでCD4およびCD8 T細胞を枯渇させるために、マウスに200 mgの抗CD4 mAb(クローンGK1.5)および200 mgの抗CD8 mAb(クローン2.43)14をi.p. 注入した。T細胞の継続的な減少を達成するために、最初の注射の2日後に、抗CD4 mAb 100 mgと抗CD8 mAb 100 mgを必要に応じて7日間の間隔で繰り返しi.p.投与した。IFN-γ中和のために、マウスに、200μlのウサギ抗ミューリンIFN-γ血清もしくは正常対照ウサギ血清、または500μgのモノクローナル抗IFN-γ抗体(Clone XMG1.2) (Bio X Cell) もしくはアイソタイプ対照mAbをBCG免疫後2週でi.p. 注入した。この処理を5dごとに1回、合計3週間繰り返した。
PKH26-PCLによるin vivo肺胞マクロファージ標識化
AMの選択的標識のために、PKH26-食細胞リンカー(PCL;Sigma-Aldrich)を製造者の指示に従ってDiluent B(20 mM)で希釈し、BCG免疫の1日前にマウスの肺に経鼻的に注入した(マウスあたり50 μl)42。コントロールマウスには、コントロールとしてPBSを投与した。翌日、PKH26またはPBSを投与したマウスのうち1匹を犠牲にし、AMの選択的標識化を確実に行った。
In vivo腸管透過性測定
トレーサーとしてFITC標識デキストラン(4 kDa; Sigma-Aldrich)を用い、in vivoでの腸管透過性を評価した57。
In vivo代謝物投与
BCG 免疫マウスで同定された明確な代謝物をマウスに投与するため、40mM 酪酸ナトリウムおよび 1.25mg ml-1L-carnitine( 共に Sigma-Aldrich 製)を DW に 3 週間添加した49。その後、AMと単球の検査の前に、マウスに代謝物を含まないDWを1週間供給した。代謝物を添加した水を供給されたマウスが同等の水分摂取量を確保するために、2日ごとに水分摂取量をモニターした。
AMのアダプティブトランスファー
5週齢の皮下BCG免疫マウスおよび非免疫マウスからBALで得たAMを、気管内ルートで移植した14。移植の4時間後にマウスにM. tuberculosisをチャレンジさせた。
盲腸の微生物移植
BCG免疫マウスおよび非免疫マウスから採取した盲腸を液体窒素中でスナップ冷凍し、使用時まで-80℃で保存した58。盲腸微生物移植の前に、マウスには、バンコマイシン、ネオマイシン、アンピシリン、メトロニダゾールをそれぞれ0.5 g l-1濃度で含む広域抗生物質カクテル(Sigma-Aldrich)を無菌DW(ad libitum)中で10日間投与し、抗生物質投与中、体重は毎日モニターされた。10mlのPBSで希釈し、37℃で30分間インキュベートした後、移植用糞便を調製した。抗生物質投与終了後、新たに調製した200μlの糞便を1日1回3日間経口投与し、5週間微生物相をコロニー化させた。
組織学的解析および顕微鏡検査
BCG投与後の大腸の組織学的変化を評価するために、組織切片をH&Eで染色した。大腸のバリア機能を評価するために、タイトジャンクション成分であるZO-1およびオクルディンを免疫組織化学で可視化した。簡単に言えば、脱パラフィン、再水和、抗原再取得後、組織切片を3%過酸化水素で10分間処理し、内因性ペルオキシダーゼ活性を阻害した。次に組織切片を5%ヤギ血清(Sigma)でブロックし、ZO-1ポリクローナル抗体(1:50)(Invitrogen Life Technologies)またはオクルディンウサギポリクローナル抗体(1:50)(Proteintech Group)と4℃にて一晩インキュベートした。シグナルは、HRP-DAB酵素染色レポーターシステム(Dako EnVision+system HRP labeled polymer anti-rabbit from Dako and DAB from Sigma-Aldrich) を用いて可視化した。結腸上皮表面で産生されたムチンを評価するために、McMaster大学医学部のMIRC Core組織学施設において、Leica Bond Rx自動染色機(Leica Biosystems)で組織切片をMUC2用に染色した。簡単に言えば、組織切片をLeica Epitope retrieval 2で前処理した後、ウサギモノクローナル(EPR23479-47)抗MUC2(Abcam)(1:3000)およびLeica Bond Refine検出キットを用いて染色を行った。組織学的検査とスコアリングは、治療群に対して盲検化された2人の研究者によって独立に検証された。
トレーニング、休息、刺激後の細胞の形態は、明視野顕微鏡-EVOS細胞イメージングシステム(Thermo Fisher Scientific)により調査し、倍率5倍および20倍で写真を撮影した。代表的な顕微鏡写真の画像は、AxioVisionデジタル画像ソフトウェア(Carl Zeiss Microscopy GmbH)を使用して、Zeiss Axio Imager 2 Research Microscopeの下で撮影されました。
AMの代謝アッセイ
AMのリアルタイム細胞代謝は、Seahorse XF Glycolysis Stress Test Kit(Agilent Technologies)を用いて、メーカーの説明書に従って、既述のように測定した14。解糖に相当する細胞外酸 化速度(ECAR)およびミトコンドリア呼吸に相当する酸素消費速度(酸化的リン酸化) は、Seahorse XFe24 Analyser(Agilent Technologies)を使用して評価した。解糖は、10-mMグルコース添加後のECARで表した。解糖能は、1-μMオリゴマイシン添加後の最大ECARで表した。解糖予備能は、解糖能と解糖の差で表した。データはWave Desktopソフトウェアバージョン2.6(Agilent Technologies)を用いて解析し、タンパク質に正規化した。
RNA単離とRNA-Seq
AMはBALを介して入手した。配列決定に十分なRNAを確保するため、1サンプルにつき2匹のマウスをプールした。グループ/条件ごとに3重のサンプルを設定した。非刺激サンプルと刺激サンプルは対にした。単離AMを1.6μg ml-1のM. tuberculosis WCLで刺激して、または刺激せずに培養した。12時間のインキュベーション後、RNase-free DNaseキットを含むRNeasy miniキット(QIAGEN)を用いて、製造者の説明書に従って、全細胞RNAを単離した。RNAサンプルは、使用するまで-80℃で保存した。RNA の品質を確認し、その後の RNA シーケンシングは McMaster University の Farncombe Metagenomic Facility で行った。RNA の完全性は Agilent バイオアナライザーを用いて確認した。すべての RNA サンプルが RIN (RNA インテグリティナンバー) 7.0 以上であることを確認し、最高品質のライブラリーを作製した。次に、RNAをポリAビーズ濃縮(NEBNext_PolyA_mRNA)工程にかけ、ポリAテールを持つmRNAを濃縮し、劣化した転写物がこの時点で隔離されないため、高品質のRNAが得られることを確認した。この工程では、リボソームRNAの除去も確実に行われた。単離したmRNAをcDNAに変換し、ライゲーションベースのライブラリプレップキット(NEBNext_Ultra_II_Directional_RNA)を用いてアダプターおよびユニークインデックスを含むライブラリとした。ライブラリーは、バイオアナライザーでサイズと分布を確認し、qPCRで濃度を確認した。その後、ライブラリーをプールし、Rapid Modeでオンボードクラスタリングを使用してIllumina HiSeq 1500の2レーン上で実行した。さらに、1サンプルあたり25Mのクラスタが得られた。
16sRNAのディープシーケンス解析による細菌プロファイリング
盲腸を無菌的に採取し、16S rRNA遺伝子のV34領域をPCRで増幅した59,60。合計50 ngの抽出DNA/サンプルを鋳型として、1UのTaq、1×バッファー、1.5 mM MgCl2, 0.4 mg ml-1 BSA, 0.2 mM dNTPs、341F (CCTACGGNGGCWGCAG) と806R (GGACTACNVGGTWTCTAAT) Illumina適応プライマーを各5 pmol使用しました。反応は94℃5分、94℃30秒、47℃30秒、72℃40秒を5サイクル、その後94℃30秒、50℃30秒、72℃40秒を25サイクル行い、最後に72℃で10分間伸長させた。得られたPCR産物は1.5%アガロースゲルで可視化した。陽性アンプリコンはSequalPrep Normalization kit (Thermo Fisher Scientific) を用いて正規化し、McMaster Genomics FacilityのIllumina MiSeqプラットフォームで塩基配列を決定した。
代謝プロファイリング
キャピラリー電気泳動と飛行時間型質量分析計(CE-TOF-MS)の組み合わせによる代謝プロファイリング。すべての化学標準物質およびキャリブレートは、Sigma-Aldrich から購入した。ブチルヒドロキシル化トルエン (BH4)、脂肪酸、ギ酸、有機酸、水酸化ナトリウム、回収/内部標準-4-フルオロ-L-フェニルアラニン (F-Phe), 3-クロロ-L-チロシン (Cl-Tyr) および D-glucose-13C6 (13C-glucose) が含まれます。アセトニトリル、イソプロパノール、メタノールおよび水を含むすべてのLC-MSグレードの溶媒は、Caledon Laboratoriesから入手した。すべての分析対象物の標準溶液は、LC-MSグレードの水でのストック溶液(50 mM)の連続希釈により調製し、冷蔵(4 ℃)で保管した。
サンプル調製に使用したPall Nanosep Omega 3-kDa ultrafiltration tubes (VWR International) は、まず脱イオン水でプレリンスし、残留添加物やバックグラウンドの汚染を除去した。チューブおよびフィルターを500 µlの脱イオン水ですすぎ、その後Eppendorf 5430(VWR International)を用いて10,000gで5分間遠心分離し、残留水分を除去した。40 µM の F-Phe と Cl-Try、および 2 mM 13C-glucose を含む 70:30 (MeOH:H2O) の混合液を氷上で予冷し、代謝物定量のための内部標準または回収標準として使用した。
すべての凍結盲腸、結腸および血清サンプルは、サンプル前処理工程の前に氷上でゆっくりと解凍した。40 µM の標準物質を含む上記の混合物 100 µl を、前処理した盲腸試料を含むチューブに加えました。その後、室温、3,000 rpm で 10 分間ボルテックスした。その後、盲腸試料を室温で10,000g、15分間遠心分離し、上清を保存した。上記ステップを繰り返し、両ステップの上清を合流させた。その後、上清をあらかじめ洗浄した限外ろ過チューブでろ過し、10,000g、15分間でろ過した。大腸サンプルは盲腸と同様のサンプル前処理を行ったが、限外ろ過のステップは除いた。血清サンプルは4倍に希釈した。解凍した25μlの血清サンプルに、200μMのF-Phe、Cl-Try、2mMの13C-グルコースを水に溶解した内部標準混合物20μlと水55μlを添加した。得られた混合物を室温で5秒間、3,000 rpmでボルテックスした。その後、混合物をあらかじめ洗浄した限外ろ過チューブに移し、10,000 gで15分間遠心分離した。抽出されたサンプルはすべて分析まで-80℃で凍結し、20 µlのアリコートをCE-TOF-MS分析用のポリプロピレンバイアルに移した61。肺サンプルは、120 μl のクロロホルムと 5 μM の回収標準物質 (F-phenylalanine, choline-d9) を含む 200 μl の混合物 (50:50, methanol:water) を、凍結乾燥肺サンプル ~5 mg を含むバイアルに加えることで処理されました。室温、3000rpmで15分間振盪した後、サンプルを3000g、4℃で15分間遠心分離してバイアル底部のタンパク質を沈殿させ、その後、二相のクロロホルムと水/メタノール(上)層を形成させた。上層の水層から一定量(150μl)を新しいバイアルに採取した。上記を繰り返し、上層水溶液を新たなバイアルに採取した。次に、結合した上部水層を、室温で窒素ガスの穏やかな流れの下で乾燥させた。肺抽出物は-80℃で保存し、分析前に40μMのクロロチロシンおよびナフタレンモノスルホン酸(内部標準)を含む50μlの水/メタノール(70:30)に再溶解させた。
血清サンプル中のLPSの測定
Thermo Fisher Scientific社製のPierce Chromogenic Endotoxin Quant Kitを使用して、血清サンプル中のLPS濃度を測定した。
細菌群集の機能プロファイルの予測
PICRUSt2を用いて、各サンプルの機能的メタゲノム内容を推定した(unrarefied OTU table)。品質保証のため、Nearest Sequenced Taxon Indexのスコアを調べたところ、すべてのサンプルで<0.15であった。PICRUSt2から得られたKO(KEGGパスウェイマップのK識別子)メタゲノム非階層化予測値を用いて、実験グループ間の予測メタゲノム存在量の差を解析した。11サンプル中10サンプル以上で予測される存在量が0であったメタゲノムについては、さらなる解析から除外した。また、少なくとも8つのサンプルで予測される存在量が10であるメタゲノムのみを、さらなる解析に含めた。存在量解析は、limma-voom approach62 に基づき、limma パッケージを使用して実施した。具体的には、アバンダンス値を丸め、VOOM変換とTMM正規化を行ってから差分解析を行った。調整後P値<0.05を示すメタゲノムについては、グループ間で有意に制御されているとみなし、KEGG Orthologyデータベース(https://www.genome.jp/kegg/ko.html)を用いて、BRITE階層からの用語を含むデータベースの第2および第3レベル、ならびにパスウェイを用いてこれらの用語を注釈し分類した。
定量化および統計解析
nの正確な値、中心の定義、分散と精度の測定、統計的有意性を含む統計パラメータは、図1-8に報告されている。同一サンプルは繰り返し測定せず、解析から除外したデータポイントもない。すべての解析はGraphPad Prismソフトウェア(バージョン9.3.1、GraphPad Software)を用いて行った。統計解析の信頼区間は95%に設定された。サンプルサイズを事前に決定するための統計的手法は用いなかったが、我々のサンプルサイズは過去の論文で報告されたものと同様である10,15,29。データの分布は正規分布と仮定したが、正式な検証は行わなかった。組織学的解析を除き、データの収集と解析は、実験条件に対してブラインドで行わなかった。
RNAseq解析では、リードを品質でフィルタリングし(少なくとも90%の塩基が品質スコア20以上でなければならない)、次に残りのリードをHISAT2を用いてmm10(UCSC)リファレンスとアライメントした。次に、HTSeq count を用いてリードをカウントした。各群の30%以上のサンプルでカウント数が10未満の遺伝子は、RのEdgeRパッケージのfilterByExpr関数で除去し、11,697個の遺伝子を得ました。これらの残りの遺伝子のカウントをTMM正規化法で正規化し、voom変換で変換した。P値は多重検定のためのBH補正63で補正し、<0.05の補正値を有意とみなした。PCAを行い、その結果をRのrglパッケージ(https://cran.r-project.org/web/package/rgl/index.html)を用いて可視化した。ヒートマップはRのgplotsパッケージ(https://cran.r-project.org/web/packages/gplots/index.html)、Volcanoプロットはlimmaパッケージを用いて行った。
マイクロバイオーム解析のために、DADA2を用いてリードを処理した。まず、Cutadaptを用いて、quality scoreが30以上、read lengthが100 bp以上のraw readsからadaptor配列とPCR primerをフィルタリングしてトリミングした(ref.64)。次に、16Sアンプリコンデータからの正確なサンプル推定パイプラインであるDADA2を用いて、トリミングされたraw readから配列バリアントを分離した。DNA配列リードは、イルミナランごとに個別にリードの品質に基づいてフィルタリングおよびトリミングされ、エラーレートが学習され、DADA2によって配列バリアントが決定された。配列バリアントテーブルは、別々のIlluminaランからのすべての情報を結合するためにマージされた。バイメラを除去し、SILVAデータベースバージョン1.3.2を用いて分類を割り当てた。最も豊富な細菌分類は、属レベルで認識された。マイクロバイオームの包括的な統計解析は、ウェブベースのプラットフォームMicrobiomeAnalyst(www.microbiomeanalyst.ca)65 を用いて実施された。データのフィルタリングは、すべてのゼロを含む、または1つのサンプルにのみ現れる特徴を分析から除外するという基準で行われた。データは総和スケール(カウントデータを各サンプルのリードの総数で割ったもの)。群集の多様性は、"α-diversity "と "β-diversity "としてプロファイリングされた。DESeq2を用いて、属レベルの操作的分類単位の存在量の差分を同定した。統計的有意水準は5%とした。P値を算出し、FDRで調整した。
代謝物解析のために、生のCE-TOF-MSデータ(dフォーマット)は、MassHunter Workstation Qualitative Analysisソフトウェア(バージョンB.06.00、Agilent Technologies、2012)を使用して処理されました。MassHunter Molecular Feature Extractor、Molecular Formula Generator ツール、および社内化合物データベースを使用して、生データから検出可能なすべての分子フィーチャーの包括的な調査を実施しました。分子特性は対称的な10 ppmのマスウィンドウを使用して抽出され、すべてのイオンはその正確な質量 (m/z)、内部標準 (Cl-Tyr) に対して正規化した相対移動時間 (RMT) およびイオン化モード (p, positive; n, negative) によって注釈が付けられました。RMTは、親化合物と同一のRMTを示す冗長な付加体やフラグメントイオンピークを除外するために使用される重要なパラメータであるため、報告されています。ピーク積分の前に、2次/3次Savitzky-Golay関数(7点)を用いてピークを平滑化した。すべての分子フィーチャーのピーク面積と移動時間をExcelワークシート(Microsoft Office)に転送し、固有の分子フィーチャーごとに相対ピーク面積(RPA)を.csvファイルとして保存した。RPAはすべての統計解析に使用された。パスウェイ解析(標的)およびPLS-DAを含む多変量データ解析は、Metaboanalyst 5.0 (www.metaboanalyst.ca)を用いて行った。すべての場合において、欠損値はデフォルト設定(検出された最低値の5分の1)で置き換えられ、メタボロームデータセットは特に断らない限り(一般化)対数変換およびオートスケーリングされた。
報告書の概要
研究デザインに関する詳細な情報は、この記事にリンクされているNature Portfolio Reporting Summaryでご覧いただけます。
データの利用可能性
RNA-seqデータはNCBIのGEOにアクセッションコードGSE213343https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE213343 で寄託されている。アデノウイルス(Ad)ワクチン接種マウスのRNAマイクロアレイ解析は既に発表されており14、アクセッションコードGSE118512でアクセスできる。マイクロバイオームシーケンスデータおよびメタボロームデータは、Mendeley Data(https://doi.org/10.17632/bvvfvz67z6)から入手可能である。
参考文献
Bekkering, S., Domínguez-Andrés, J., Joosten, L. A. B., Riksen, N. P. & Netea, M. G. Trained immunity: reprogramming innate immunity in health and disease(訓練された免疫:健康と病気における自然免疫の再プログラム). Annu. Rev. Immunol. 39, 667-693 (2021).
論文
CAS
Google Scholar
Netea, M. G. et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 20, 375-388 (2020).
論文
CAS
Google Scholar
Xing, Z. et al. 組織常在マクロファージの自然免疫記憶と訓練された自然免疫:ワクチンコンセプトと戦略の再構築. J. Leukoc. Biol. 108, 825-834 (2020).
論文
CAS
Google Scholar
Benn, C. S., Netea, M. G., Selin, L. K. & Aaby, P. A small jab-a big effect: nonspecific immunomodulation by vaccines.(ワクチンによる非特異的免疫調節)。Trends Immunol. 34, 431-439 (2013).
論文
CAS
Google Scholar
Giamarellos-Bourboulis, E. J. et al. Activate: randomized clinical trial of BCG vaccination against infection in the elderly.(アクティベート:高齢者におけるBCGワクチン接種の無作為化臨床試験)。Cell 183, 315-323 (2020).
論文
CAS
Google Scholar
Prentice, S. et al. BCGによるウガンダ新生児の異種感染症に対する非特異的効果:治験責任医師盲検化無作為化比較試験. Lancet Infect. Dis. 21, 993-1003 (2021).
論文
CAS
Google Scholar
Khader, S. A. et al.結核ワクチン接種のための自然免疫の標的化。J. Clin. Invest. 129, 3482-3491 (2019).
論文
Google Scholar
Mitroulis, I. et al. 骨髄造血前駆細胞の調節は、訓練された免疫の不可欠な構成要素である。Cell 172, 147-161 (2018)。
論文
キャス
Google Scholar
Cirovic, B. et al.ヒトにおけるBCGワクチン接種は、造血前駆細胞コンパートメントを介して訓練された免疫を誘発する。Cell Host Microbe 28, 322-334 (2020).
論文
CAS
Google Scholar
Moorlag, S. J. C. F. M. et al. β-グルカンは結核菌感染に対する保護的な訓練された免疫を誘導する:IL-1の重要な役割. Cell Rep. 31, 107634 (2020).
論文
CAS
Google Scholar
Cunningham, K. T., Finlay, C. M. & Mills, K. H. G. Helminth imprinting of hematopoietic stem cells sustains anti-inflammatory trained innate immunity that attenuates autoimmune disease(造血幹細胞の蠕虫インプリンティングは、自己免疫疾患を抑制する自然免疫を維持する。J. Immunol. 206, 1618-1630 (2021).
論文
CAS
Google Scholar
Christ, A. et al. Western diet triggers NLRP3-dependent innate immune reprogramming. Cell 172, 162-175 (2018)です。
論文
CAS
Google Scholar
Kaufmann, E. et al. BCGは造血幹細胞を教育し、結核に対する保護的な自然免疫を生成させる。Cell 172, 176-190 (2018).
論文
キャス
Google Scholar
Yao, Y. et al. Induction of autonomous memory alveolar macrophages requires T cell help and is critical to trained immunity. Cell 175, 1634-1650 (2018年)。
論文
CAS
Google Scholar
Guillon, A. Pneumonia recovery reprograms the alveolar macrophage pool.肺炎の回復が肺胞マクロファージプールを再プログラムする。JCI Insight 5, e133042 (2020).
論文
Google Scholar
Machiels, B. et al. A gammaherpesvirus provides protection against allergic asthma by inducing the replacement of resident alveolar macrophage with regulatory monocytes.(ガンマヘルペスウィルスは肺胞マクロファージの制御性単球への置換を誘導することにより、アレルギー性喘息に対する保護を提供する。Nat. Immunol. 18, 1310-1320 (2017).
論文
キャス
Google Scholar
Roquilly, A. et al. 肺胞マクロファージは炎症後にエピジェネティックに変化し、長期の肺免疫麻痺につながる。Nat. Immunol. 21, 636-648 (2020).
論文
CAS
Google Scholar
Roquilly, A. et al. 肺炎治癒後の抗原提示細胞発達の局所的調節は、二次感染に対する長期的な感受性を誘導する。Immunity 47, 135-147 (2017).
論文
キャス
Google Scholar
D'Agostino, M. R. et al. Airway macrophage mediate mucosal vaccine-induced trained innate immunity against Mycobacterium tuberculosis in early stages of infection.気道マクロファージは、結核菌に対する粘膜ワクチンによる訓練された自然免疫を媒介する。J. Immunol. 205, 2750-2762 (2020).
論文
Google Scholar
肺における訓練された免疫と自然免疫の局所的な記憶。Cell 175, 1463-1465 (2018).
論文
キャス
Google Scholar
Guilliams, M. & Svedberg, F. R.組織インプリンティングはマクロファージの可塑性を制限するか?Nat. Immunol. 22, 118-127 (2021).
論文
CAS
Google Scholar
Guilliams, M., Thierry, G. R., Bonnardel, J. & Bajenoff, M. マクロファージニッチの成立と維持。Immunity 52, 434-451 (2020).
論文
CAS
Google Scholar
オルドバス-モンタネス, J., ベヤズ, S., ラコフ-ナホウム, S. & シャレック, A. K. バリア組織における炎症性記憶の分布と保存. Nat. Rev. Immunol. 20, 308-320 (2020).
論文
CAS
Google Scholar
Kadoki, M. et al. Organism-level analysis of vaccination reveal networks of protection across tissues. Cell 171, 398-413 (2017)。
論文
CAS
Google Scholar
Krausgruber, T. et al. Structural cells are key regulators of organ-specific immune responses(構造細胞は臓器特異的免疫応答の重要な制御因子である)。ネイチャー 583, 296-302 (2020)。
論文
CAS
Google Scholar
Hoyer, F. F. et al. 遠隔傷害に対する組織特異的マクロファージ応答は、その後の局所免疫チャレンジの結果に影響を与える。Immunity 51, 899-914 (2019).
論文
キャス
グーグル スカラー
Budden, K. F. et al. Emerging pathogenic links between microbiota and the gut-lung axis(マイクロバイオータと腸-肺軸の間の新たな病原性関連性)。Nat. Rev. Microbiol. 15, 55-63 (2017).
論文
キャス
Google Scholar
Dang, A. T. & Marsland, B. J. Microbes, metabolites, and the gut-lung axis. Mucosal Immunol. 12, 843-850 (2019).
論文
キャス
Google Scholar
Trompette, A. et al. Dietary fiber confers protection against flu by shaping Ly6c- patrolling monocyte hematopoiesis and CD8+T cell metabolism.食物繊維は、Ly6c-パトローリング単球の造血とCD8+T細胞の代謝を形成することによって、インフルエンザに対する保護を与える。Immunity 48, 992-1005 (2018).
論文
キャス
Google Scholar
Sencio, V. et al. インフルエンザ時の腸内細菌異常は、短鎖脂肪酸産生の変化を通じて肺炎球菌のスーパーインフェクションに寄与する. Cell Rep. 30, 2934-2947 (2020).
論文
CAS
Google Scholar
Khan, N. et al.腸内細菌異常は結核菌に対する肺胞マクロファージ免疫を損なう。Mucosal Immunol. 12, 772-783 (2019).
論文
キャス
Google Scholar
Samuelson, D. R. et al. Alcohol-associated intestinal dysbiosis impairs pulmonary host defense against Klebsiella pneumoniae.アルコールに関連する腸内細菌症は、肺炎菌に対する肺の宿主防御を損ないます。PLoS Pathog. 13, e1006426 (2017).
論文
Google Scholar
Trompette, A. et al. 腸内細菌叢による食物繊維の代謝は、アレルギー性気道疾患および造血に影響を及ぼす。Nat. Med. 20, 159-166 (2014).
論文
CAS
Google Scholar
Rosshart, S. P. et al. Wild mouse gut microbiota promotes host fitness and improves disease resistance(野生のマウス腸内細菌叢は宿主の体力を高め、病気への抵抗力を向上させる。Cell 171, 1015-1028 (2017)。
論文
キャス
Google Scholar
Jeyanathan, M., Yao, Y., Afkhami, S., Smaill, F. & Xing, Z. New tuberculosis vaccine strategies: taking aim at un-natural immunity.(新しい結核ワクチン戦略:非自然免疫に狙いを定める)。Trends Immunol. 39, 419-433 (2018).
論文
CAS
Google Scholar
Jeyanathan, M. et al. Immunological Considerations for COVID-19 vaccine strategies. Nat. Rev. Immunol. 20, 615-632 (2020).
論文
CAS
Google Scholar
Louwe, P. A. et al. 炎症後の腹膜ニッチをコロニー化するリクルートマクロファージは、機能的に多様な常在細胞に変化する。Nat. Commun. 12, 1770 (2021).
論文
CAS
Google Scholar
Cohen, S. B. et al. Alveolar macrophage provides an early mycobacterium tuberculosis niche and initiate dissemination(肺胞マクロファージは初期の結核菌ニッチを提供し、播種を開始する。Cell Host Microbe 24, 439-446 (2018).
論文
キャス
Google Scholar
Lai, R. et al. CD11b+樹状細胞を介した抗結核菌Th1活性化は、IL-10を介してCD103+樹状細胞によって逆調節される。J. Immunol. 200, 1746-1760 (2018).
キャス
Google Scholar
Jeyanathan, M. et al. CXCR3シグナルは、肺実質と気道の両方への非経口結核ワクチン誘導T細胞の制限されたホーミングに必要である。J. Immunol. 199, 2555-2569 (2017).
論文
キャス
Google Scholar
Delahaye, J. L. et al. Cutting edge: Bacillus Calmette-Guérin-induced T cells shape Mycobacterium tuberculosis infection before reducing the bacterial burden.(バチルスカルメットゲリン誘発T細胞は、細菌の負担を減らす前に結核菌の感染を形成する。J. Immunol. 203, 807-812 (2019).
論文
キャス
Google Scholar
Califano, D., Furuya, Y. & Metzger, D. W. Effects of influenza on alveolar macrophage viability are dependent on mouse genetic strain.インフルエンザの肺胞マクロファージ生存率への影響は、マウスの遺伝子系統に依存する。J. Immunol. 201, 134-144 (2018).
論文
キャス
Google Scholar
Erben, U. et al. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 7, 4557-4576 (2014).
Google Scholar
Douglas, G. M. et al. PICRUSt2 for prediction of metagenome functions(PICRUSt2によるメタゲノム機能の予測)。Nat. Biotechnol. 38, 685-688 (2020).
論文
CAS
Google Scholar
GATA6を持つ腹腔マクロファージは腸漿膜損傷の修復に寄与する。Nat. Commun. 12, 7294 (2021).
論文
CAS
Google Scholar
Wang, J. & Kubes, P. A reservoir of mature cavity macrophage that can rapidly invade visceral organs to affect tissue repair.(内臓臓器に急速に侵入して組織修復に影響を及ぼす成熟した空洞マクロファージのリザーバー)。Cell 165, 668-678 (2016)です。
論文
CAS
Google Scholar
Yan, H., Baldridge, M. T. & King, K. Y. Hematopoiesis and the bacterial microbiome(造血と細菌マイクロバイオーム). Blood 132, 559-564 (2018)。
論文
CAS
Google Scholar
Ifrim, D. C. et al. Trained immunity or tolerance: Opposing functional programs induced in human monocytes after engagement of various pattern recognition receptors.(訓練された免疫または寛容:様々なパターン認識受容体の関与後にヒト単球に誘導される相反する機能プログラム)。Clin. Vaccine Immunol. 21, 534-545 (2014).
論文
Google Scholar
Erny, D. et al. Host microbiota constantly control maturation and function of microglia in the CNS.(宿主微生物叢は、CNSにおけるミクログリアの成熟と機能を絶えず制御している。Nat. Neurosci. 18, 965-977 (2015).
論文
キャス
Google Scholar
Silva, F. et al. マウス皮下BCGワクチン接種と結核菌感染により、肺と腸の微生物叢が変化する。Microbiol. Spectr. 10, e0169321 (2022).
論文
Google Scholar
Huda, M.N.ら、乳児期早期のビフィズス菌の存在量と2歳時のワクチン反応。Pediatrics 143, e20181489 (2019).
論文
Google Scholar
Koeken, V. A. C. M., Verrall, A. J., Netea, M. G., Hill, P. C. & van Crevel, R. Trained innate immunity and resistance to Mycobacterium tuberculosis infection.(結核菌感染症に対する自然免疫の訓練と耐性). Clin. Microbiol. Infect. 25, 1468-1472 (2019).
論文
キャス
Google Scholar
Neupane, A. S. et al. Patrolling alveolar macrophage conceal bacteria from the immune system to maintain homeostasis.(パトロールする肺胞マクロファージは免疫システムから細菌を隠し、恒常性を維持する)。細胞 183, 110-125 (2020)。
論文
CAS
Google Scholar
非経口BCGワクチン接種者の肺における抗結核防御遅延のメカニズム:気道内腔T細胞の重要な役割。Mucosal Immunol. 5, 420-431 (2012).
論文
CAS
Google Scholar
Smith, S. G., Kleinnijenhuis, J., Netea, M. G. & Dockrell, H. M. 全血プロファイリングにより、乳児におけるバシラスカルメットゲリン誘発訓練自然免疫の上皮成長因子、IL-6、プレートレット由来成長因子AB/BBおよびナチュラルキラー細胞の活性化を特定した。Front. Immunol. 8, 644 (2017).
論文
Google Scholar
& Jeyanathan, M. et al. 粘膜免疫によるマウス気道管腔抗結核メモリーCD8 T細胞は、末梢T細胞のリクルートとは無関係に、抗原駆動型のin situ増殖を介して維持される。Am. J. Respir. Crit. Care Med. 181, 862-872 (2010).
論文
CAS
Google Scholar
Thevaranjan, N. et al. Age-associated microbial dysbiosis promotes intestinal permeability, systemic inflammation, and macrophage dysfunction.加齢に伴う微生物ディスバイオーシスは、腸管透過性、全身性炎症、およびマクロファージ機能不全を促進する。Cell Host Microbe 21, 455-466 (2017).
論文
キャス
Google Scholar
Kwon, Y. H. et al. セロトニンシグナルによる腸内細菌叢組成の調節は、腸管免疫反応および大腸炎に対する感受性に影響を与える。Cell. Mol. Gastroenterol. Hepatol. 7, 709-728 (2019).
論文
Google Scholar
Stearns, J. C. et al. Culture and molecular-based profiles show shifts in bacterial communities of the upper respiratory tract that occur with age.(培養および分子ベースのプロファイルは、年齢とともに生じる上気道の細菌群集のシフトを示す)。ISME J. 9, 1246-1259 (2015).
論文
Google Scholar
Bartram, A. K., Lynch, M. D. J., Stearns, J. C., Moreno-Hagelsieb, G. & Neufeld, J. D. Generation of multimillion-sequence 16S rRNA gene libraries from complex microbial communities by assembling paired-end illumina reads.(イルミナリードのペアエンドによる複雑な微生物群集からの数百万シーケンス16S rRNA遺伝子ライブラリの生成)。Appl. Environ. Microbiol. 77, 3846-3852 (2011).
論文
CAS
Google Scholar
Shanmuganathan, M. et al. Multisegment injection-capillary electrophoresis-mass spectrometry by maternal serum metabolome: a high-throughput platform and standardized data workflow for large-scale epidemiological studies.による母体血清メタボロームの解析。Nat. Protoc. 16, 1966-1994 (2021).
論文
CAS
Google Scholar
Ritchie, M. E. et al. Limma powers differential expression analyses for RNA-sequencing and microarray studies.リッチー、M.E.ら、RNAシーケンスおよびマイクロアレイ研究のための差分発現解析。Nucleic Acids Res. 43, e47 (2015).
論文
Google Scholar
Benjamini, Y. & Hochberg, Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing.ベンジャミニ、Y.およびホッホベルク、Y.は、偽発見率を制御する。J. R. Stat. Soc. B 57, 289-300 (1995).
Google Scholar
Kechin, A., Boyarskikh, U., Kel, A. & Filipenko, M. cutPrimers: a new tool for accurate cutting of primers from reads of targeted next generation sequencing.「次世代シーケンサーのリードからプライマーを正確にカットするツール」。J. Comput. Biol. 24, 1138-1143 (2017)に掲載されています。
論文
CAS
Google Scholar
Chong, J., Liu, P., Zhou, G. & Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data.(マイクロバイオームデータの包括的統計解析、機能解析、メタ解析のためのMicrobiomeAnalystの使用)。Nat. Protoc. 15, 799-821 (2020).
論文
CAS
グーグルスカラー
参考文献のダウンロード
謝辞
本研究は、カナダ保健研究所(CIHR)基金プログラム(FDN-154316 to Z.X.)、カナダ国立療養所協会(to Z.X.)、カナダイノベーション基金、ゲノムカナダ(to P.B.-M. )、オンタリオ政府およびマクマスター大学からの資金援助により行われました。M. TarnopolskyにはSeahorse XFe24アナライザーの使用を許可していただき、Z. HmamaにはBCG-dsRed株の提供をお願いした。
著者情報
著者および所属
McMaster Immunology Research Centre, M. G. DeGroote Institute for Infectious Disease Research and Department of Medicine, McMaster University, Hamilton, Ontario, Canada
Mangalakumari Jeyanathan, Maryam Vaseghi-Shanjani, Sam Afkhami, Alisha Kang, Michael R. D'Agostino, Yushi Yao, Shreya Jain, Anna Zganiacz, Ramandeep Singh, Anna Dvorkin-Gheva & Zhou Xing
ファーンコム・ファミリー消化器健康研究所およびマクマスター大学病理学・分子医学部(カナダ、オンタリオ州、ハミルトン市
Jensine A. Grondin & Waliul I. Khan
浙江大学免疫学教室(中国・浙江省
ヤオ・ユシ
マクマスター大学化学・ケミカルバイオロジー学科(カナダ・オンタリオ州ハミルトン市
ザッカリー・クローゼン、ミーラ・シャンムガナサン、フィリップ・ブリッツ・マッキビン
寄稿
M.J.とZ.X.は本研究を構想し、設計した。M.J., M.V.-S., S.A., J.A.G., A.K., M.R.D., Y.Y., S.J., A.Z., Z.K., M.S. and R.S.は実験を実施した。M.J.、M.V.-S.、M.S.、A.D.-G.はデータを解析した。P.B.-M.とW.I.K.は代謝とマイクロバイオミクスの研究を監修した。M.J.とZ.X.は論文を執筆した。
共著者
Zhou Xingにご連絡ください。
倫理的宣言
利益相反
著者らは、競合する利益を宣言していない。
査読
査読情報
Nature Immunology誌は、Antoine Roquilly氏と他の匿名の査読者の方々に感謝します。主な取扱編集者 Nick Bernard、Nature Immunologyチームと共同。査読者のレポートが利用可能です。
その他の情報
出版社からのコメント Springer Natureは、出版された地図や所属機関に関する管轄権の主張に関して中立的な立場を維持しています。
エクステンデッドデータ
Extended Data 図1 BCG s.c. ワクチン接種後の肺胞マクロファージの自然免疫記憶表現型の特性評価。
(a) 刺激あり(S)または刺激なし(US)の肺組織AM上のMHC II発現のMFI(**p = 0.0012, ***p = 0.0004). (b & c) IL-6およびTNF産生肺組織AMの頻度、(S)または(US)刺激なし。(d)BCGワクチン接種後8週目の気道AMにおけるリアルタイム酸素消費率(OCR)。(e)実験スキーマ。(f & g) 気道AMによるBrdUの組み込みのヒストグラムおよびBrdUが組み込まれた気道AMの頻度 (*p = 0.0475) (f) およびBrdU + (*p = 0.0430) およびBrdU- (p = 0.0519) 気道AMにおけるMHCIIのMFI (g). n = 3 マウス/グループ. (h) 実験スキーマ。(i, j & k) 気道および肺組織AMのMHCIIのMFIの測定 (*p = 0.0365) (i), IL-6- (j) およびTNF- (k) 産生気道および肺組織AMの頻度 (*p = 0.0249) n = 3匹/グループ/気道, 4匹/グループ/肺組織。(l)気道AMのリアルタイム細胞外酸性化率(ECAR)、および気道AMの解糖、解糖能、解糖予備能(***p = 4マウス/グループ。(m)生BCG、不活化BCG(BCG-ia)またはPBSによるワクチン接種後5週間、再刺激あり(S)またはなし(US)での気道AMのMHCII発現のMFI(*p = 0.0497)およびIL-6(**p = 0.0082)およびTNF生成気道AMの頻度(n = 3マウス/グループ)の測定。a-cのデータは、2つの独立した実験の代表値である(n = 3マウス/グループ)。棒グラフのデータは、平均値±SEMとして示される。a-cとi-kのデータは一元配置分散分析にボンフェローニ検定を、f, g, lのデータは両側t検定を、mは一元配置分散分析にダネット多重比較検定を適用して多重比較を行った。
Extended Data 図2 BCGワクチン接種および呼吸器粘膜ウイルスワクチン接種によって誘導されたメモリー肺胞マクロファージの転写シグネチャー。
(a) 比較のうち少なくとも1つによって気道AMで差次的に発現した遺伝子のヒートマップ。絶対的なフォールド変化が少なくとも1.5であった場合に差とした。データは各グループの各遺伝子の平均発現レベル。(b)刺激(US)なしのBCGワクチン接種動物対PBS動物の気道AMの比較のためのボルカノプロットで、絶対的な倍数変化の閾値は1.5に設定された。(c) M.tb WCL刺激によるBCGワクチン接種動物対PBS動物の気道AMの比較のボルケーノプロット(絶対倍率変化の閾値を1.5に設定)。(d) BCGワクチン接種動物からの未刺激(US)気道AMにおいて、PBS処理動物からのものと比較して有意に発現が増加した遺伝子のパスウェイを示す。(e & f) BCG接種動物の気道AMにおける解糖(e)およびmTOR(f)に関与する遺伝子のシグネチャスコアを、対照PBS動物からのものと、刺激あり(S)または刺激なし(US)で比較したものである。(g)絶対倍数変化の閾値を1.5に設定した、刺激なし(US)のアデノウイルスベクターワクチン(Ad)経鼻免疫動物対PBS対照動物からの気道AMの比較のためのボルケーノプロット(Volcano plot)。(h) アデノウイルスワクチン(Ad)経鼻免疫動物の気道AMにおけるAg処理/提示 (****p = 6.15×10-7), 脂肪酸酸化 (*p = 1.6×10-2), 解糖およびmTOR経路に関わる遺伝子の署名スコアを、(US)刺激を行わないPBSコントロール動物からの遺伝子と比較することで、アデノウイルスワクチン(Ad)を経鼻免疫動物から得た。バイオリンプロットの横線は中央値、点線は下位および上位四分位を示す。a-gのデータは3サンプル/グループ/条件での実験から得られたものである。volcano plot (b, c & g)の縦線は絶対倍率1.5の閾値を、横線は0.05に設定した修正p値を示す。有意な遺伝子は色で示され、発現量の多い遺伝子は赤で、発現量の少ない遺伝子は青で示されている。調整後のp値はバイオリンプロット(h)で示され、limmaパッケージとBH補正を用いて得られた(Methods参照)。
Extended Data 図3 記憶肺胞マクロファージの抗原提示および抗菌能力の特性評価。
(a)実験スキーマ b)実験スキーマ (b) BCG ワクチン接種動物または PBS 処理動物の Ag85 を負荷した気道 AM に応答して増殖する Ag85B 特異的 CD4 T 細胞の代表的ヒストグラム(CFSE 標識希釈の程度として較正)。(c) 増殖の各世代 (G) におけるAg85B+ CD4 T細胞の頻度。(d) BCGワクチン接種またはPBS処理動物からの肺組織CD11C+CD11b+抗原提示細胞(APC)によってM.tb感染後24(***p(**p = 0.0006))で殺された食作用細菌のパーセンテージ、n = 4マウス/グループ。(e)感染後24時間におけるBCG-ワクチン接種動物およびPBS処理動物の壊死およびアポトーシスBCG-dsRed感染気道AMの代表的なフロープロットおよび頻度。棒グラフのデータは、平均±SEMとして示される。統計解析は、cおよびdのデータについて、両側t検定により決定した。
Extended Data 図4 肺M.tb感染後の肺組織におけるCD4 T細胞応答の動態。
(a) 表面マーカーCD4およびCD44で免疫染色した肺単核細胞の代表的ドットプロット。5週間のPBS処置またはBCGワクチン接種マウスを犠牲にする3-5分前にCD45.2抗体を静脈内投与し、真正肺実質性T細胞(LPT)と肺血管内細胞(LV)を区別するゲーテイングを行った。データは1回の実験によるものである。(b)マウスは5週間、BCG-ワクチン接種またはPBS-ワクチン接種をs.c.された。PBS処理(PBS-AM)およびBCG-ワクチン接種(BCG-AM)から気管支肺胞洗浄によって採取した気道AMをナイーブマウスに養子移入し、M.tbにチャレンジさせた。感染後3日目の肺のM.tb CFUを評価した(1群あたりn = 4マウス)。データは1回の実験から得たものである。
Extended Data 図5 循環単球の免疫特性評価。
(a) 末梢血中の循環単球の同定に使用したゲーティング戦略。CD3+T細胞(CD3-)とCD11b+Ly6G+好中球(好中球-)を除去するため、生きたCD45+細胞をゲーティングした。残りの細胞集団は、古典的なCD11b+Ly6Chighと非古典的なCD11b+Ly6Clow循環単球を同定するためにゲーティングされた。(b) BCGワクチン接種およびPBS処理した宿主の末梢血中のLy6ChighおよびLy6Clow単球上のMHCII発現レベルを、刺激あり(S)および刺激なし(US)で培養した代表的ヒストグラム(n = 3マウス/群)。(c & d) IL-6産生Ly6ChighおよびLy6Clow単球の頻度、およびTNF産生Ly6Chigh単球の頻度を、(S)および(US)刺激なしで培養した場合。(e)IL-6(*p=0.015)、IL-1β(**p=0.0007)、TNF、IL12p40(*p=0.0173)、IP10(*p=0.0229)、MIP-1-α(*p=0.0407)およびRANTES(*p=0.0024)。 0024)タンパク質含量を、(S)刺激ありおよび(US)刺激なしで培養したBCGワクチン接種またはPBS対照動物由来の全血サンプルの血漿中(pg/ml)。n = 3マウス/グループ。データは、2つの独立した実験の代表として、平均±SEMで示される。統計解析は、e.のデータについて両側t検定により決定した。
Extended Data 図6 肺のマクロファージサブセット、CCR2KOマウスの肺胞マクロファージによるex vivo M.tb殺傷、およびメモリー肺胞マクロファージの誘導におけるIFN-γの役割。
(a & b) PBS投与またはBCG接種後2週間の気道および肺組織におけるCD64+CD24-マクロファージにゲーティングしたSiglecF+Ly6C-肺胞マクロファージ(AM)の代表ドットプロット、肺組織の単球由来(MDM)と間質(IM)マクロファージをそれぞれSiglecF-Ly6C+とSiglecF-Ly6C-として識別。気道および肺組織における異なるマクロファージサブセットの総数が棒グラフに示されている。(c) 5週間のBCGワクチン接種またはPBS処理したCCR2KO動物からの気道AMのマイコバクテリア殺傷能力を、ex vivo殺傷アッセイで評価した。細胞内M.tb CFUの総数をex vivo感染後24時間目に評価した。各ポイントは生物学的に独立したサンプルを表す(*p = 0.0351)。n = 3 wells/PBS; n = 4 wells/BCG. データは、平均±SEMとして示される。(d)PBS対照、BCG-ワクチン接種またはBCG-ワクチン接種/IFNγ枯渇動物からのTNF産生気道AMの頻度(非刺激-US対刺激-S)(*p=0.0031)n=3マウス/群。データは、平均±SEMとして示される。およびdのデータは、多重比較のための一元配置分散分析、続いてボンフェローニ検定によって分析される。cのデータの統計解析は、両側t検定によって決定された。
Extended Data 図7 BCGワクチン接種後の腸内細菌叢、組織形態学、および上皮接合部タンパク質分布の変化。
(a) 対照のPBS処理動物とBCGワクチン接種動物の盲腸の異なる肉眼的外観。n = 5 マウス/グループ (b) 5 週間の BCG ワクチン接種動物(n = 5)と PBS 対照動物(n = 6)における OTU データに基づく上位 9 細菌群の相対存在量プロファイル。2つの独立した実験の代表例。(c) 2週間BCG接種動物(n = 5)と対照PBS(n = 4)間のChao1多様性測定(t検定)を用いた盲腸微生物叢の操作的分類単位(OTU)富に基づくアルファ多様性の比較。箱型プロットの横線は中央値を、箱の長さは下位四分位と上位四分位を、ひげは最小値と最大値を表す。(d) 2週間のBCGワクチン接種動物と対照PBS動物におけるOTUデータに基づく上位8細菌群の相対存在量プロファイル。(e) PCoA順序法およびJenson-Shannon発散距離法(PERMANOVA)を用いた、2週間のBCGワクチン接種動物とPBS動物における糞便微生物群集のベータ多様性の比較。(f) 2週間BCG接種した宿主の糞便微生物叢における上位4つの豊富な細菌ファミリーの頻度を、PBS対照宿主のものと比較した棒グラフ。(g)5週目のBCGワクチン接種動物およびPBS対照動物の結腸の建築的変化、上皮の変化および炎症性浸潤の組織学的スコアリング。2つの独立した実験の代表。n = 6マウス/グループ。(h)2-週、5-週、8-週のBCG-ワクチン接種宿主およびPBS対照宿主からの結腸組織の組織学的比較。BCG接種後のn = 5匹/2wk, 6匹/5wk, 4匹/8wkの代表例。(i)5週目のBCGワクチン接種動物およびPBS対照動物における腸上皮接合部オクルーディンタンパク質の免疫組織化学的染色。n=6マウス/PBS、5マウス/BCGの代表。棒グラフのデータは、平均値±SEMで示した。
Extended Data 図8 BCG接種後の腸、血清および肺における微生物代謝プロファイルの変化、および血清中のLPSが検出されないこと。
(a) 5週間のBCGワクチン接種動物とPBS動物の盲腸組織試料におけるクレアチニンレベルの比較。*p = 0.0330 (b) BCG接種5週齢動物とPBS動物の大腸組織サンプルにおける代謝プロファイルに関するPLS-DA分析。(c) BCG接種5週齢動物およびPBS動物の結腸組織サンプルにおける乳酸レベルの比較。*p = 0.0122 (d) 投影変数重要度(VIP)分析により同定されたモデル全体に対する血清代謝物の重要性。右側の色のついたボックスは、5 週間の BCG 接種を受けた血清サンプルと PBS コントロール血清サンプルの血清代謝物の比較レベルを示す。(e)Metaboanalyst 5.0、Mus musculus(KEGG)パスウェイライブラリを用いた「パスウェイ解析-ターゲット」で解析した、VIPスコア1.5以上の5週齢BCG接種動物とPBS対照動物の血清代謝物のパスウェイ解析。VIPスコア1.5以上の血清代謝物には、3つのパスウェイが有意に関与していることが確認された。(f) BCG接種5週齢動物とPBS動物の糞便組織サンプル中のSCFAsの総濃度(mM/g)のうち、酢酸およびプロピオン酸SCFAsの割合の算出。*p = 0.0234 (g) 3週および5週BCG接種動物およびPBS対照動物の血清中のLPSレベル。(h) 2週および5週のBCG接種動物およびPBS対照動物(n = 10マウス/グループ)の肺組織代謝産物を、投影における変数重要度(VIP)分析によって同定したもの。右側の色のついたボックスは、BCG接種動物と対照動物との代謝物の比較レベルを示す。a-fのデータは、2つのプールされた実験(n = 16 mice/group)から得られたものである。箱ひげ図中の横線は中央値を、箱の長さは下位四分位と上位四分位を、ひげは最小値と最大値を示している。統計解析は、a, c, fのデータについて両側t検定で決定した。
Extended Data Fig. 9 BCG接種動物における循環微生物代謝産物による肺胞マクロファージの自然免疫訓練。
(a) BCG調整血清または対照血清で訓練した後、再刺激の前に安静にしているAMの代表的な明視野顕微鏡画像。矢印は、BCG調整血清を介した訓練と細胞分裂に起因するAMのクラスターとサイズの増加および広がりを指している。(b) DWまたはDW + Mの動物からの未刺激(US)または再刺激(S)の気道AMの培養上清中のサイトカイン/ケモカインタンパク質レベル(幾何平均)のヒートマップ。アスタリスクは、DW + M宿主の気道AMによる刺激でサイトカイン/ケモカイン生産が著しく増加することを示している。(c) DW+Mの動物の循環中の古典的CD11b+Ly6Chigh単球がDWの動物に比べて減少していることを示す代表的なドットプロット。(d)DW対DW+M動物の循環単球上のLy6CまたはMHCII発現のMFI(*p = 0.0314, ***p = 0.0007)。データは平均±SEMとして示される。n=4マウス/DW、3マウス/DW+M。(e)DWおよびDW+M動物の非刺激(US)および刺激(S)全血培養の血漿中のサイトカイン/ケモカインのタンパク質レベル(幾何平均値)のヒートマップ。(f)DWおよびDW+Mの動物からのH&E染色された結腸組織切片の代表的な顕微鏡写真。 g)DW+Mの動物からの気道AMにおける選択遺伝子の発現の、刺激(US)なしのDWの動物との比較に関するボルカノプロット、絶対倍変化の閾値は、1.(h) DW-M上の動物と刺激後のDW上の動物からの気道AMにおける選択遺伝子の発現の比較のためのボルケーノプロット、絶対倍数の変化の閾値は1.5に設定される。(i-k)DW+MまたはDWを投与した動物の気道AMにおける脂肪酸酸化(*p = 0.0209)、解糖およびmTOR(*p = 0.0209)、刺激あり(S)または刺激なし(US)の遺伝子のシグネチャスコアを比較したもの。バイオリンプロットの横線は中央値、点線は下位および上位四分位を示す。a-cのデータは2つの独立した実験の代表値である(n = 4 wells/condition)。volcano plot (g & h)の縦線は絶対倍数変化の閾値1.5を、横線はp値0.05で調整したものを示す。有意な遺伝子を色で示し、発現量の多いものを赤、発現量の少ないものを青で示した。統計解析はdのデータに対して両側t検定で決定した。調整後のp値はバイオリンプロット(iとk)に対して示し、limmaパッケージとBH補正を用いて求めた(方法参照)。
Extended Data 図10 関連分野で以前に観察された内容との関連で、本研究から得られた主要な知見を示す図。
近年、ワクチンや微生物の非経口/全身曝露により、骨髄の骨髄前駆細胞に刷り込まれ、訓練された単球が血流に放出されることにより、中枢性自然免疫記憶と訓練された免疫(TII)が誘導されることが示されている。一方、ワクチンや微生物に呼吸器粘膜が暴露されると、粘膜組織常在の記憶マクロファージや肺のTIIが誘導される可能性がある。また、呼吸器感染症が腸内細菌叢の異常につながるというエビデンスもある。しかし、非経口ワクチン接種が腸内細菌叢の異常を引き起こし、腸管バリア機能やメタボロームの変化をもたらし、その結果、遠位粘膜器官である肺に粘膜組織常在型メモリーマクロファージやTIIを誘導するかどうかは不明なままであった。BCG接種後、BCG菌は腸管組織に拡散し、腸内細菌叢やバリアー透過性の時間依存的変化、メタボローム変化を腸管だけでなく血清や肺組織でも引き起こすことが明らかになった。このように、微生物のメタボローム経路を介した多くの組織部位にわたる広範な免疫学的警告は、最終的に、訓練された組織常在マクロファージと自然免疫の時間依存的誘導を肺にもたらす。
補足情報
報告書概要
ピアレビューファイル
補足表
補足表1~5
権利と許可
オープンアクセス 本論文は、クリエイティブ・コモンズ 表示 4.0 国際ライセンスの下に提供されており、原著者と出典に適切なクレジットを与え、クリエイティブ・コモンズ・ライセンスへのリンクを提供し、変更を加えたかどうかを示す限り、あらゆる媒体や形式での使用、共有、適応、配布、複製を許可しています。この記事に掲載されている画像やその他の第三者の素材は、素材へのクレジット表示で別段の指示がない限り、記事のクリエイティブ・コモンズ・ライセンスに含まれます。素材が記事のクリエイティブ・コモンズ・ライセンスに含まれておらず、あなたの意図する利用が法令上の規制で許可されていない場合、または許可された利用を超える場合には、著作権者から直接許可を得る必要があります。このライセンスのコピーを見るには、http://creativecommons.org/licenses/by/4.0/。
転載と許可
この記事について
クロスマークで通貨と真偽を確認する
この記事の引用
Jeyanathan, M., Vaseghi-Shanjani, M., Afkhami, S. et al. 非経口BCGワクチンは、腸-肺軸を介して肺内在性メモリーマクロファージと訓練免疫を誘発する。Nat Immunol 23, 1687-1702 (2022)。https://doi.org/10.1038/s41590-022-01354-4。
引用文献のダウンロード
受付終了
2021年11月02日
Accepted
2022年10月05日
掲載
2022年12月01日発行
発行日
2022年12月
DOI
https://doi.org/10.1038/s41590-022-01354-4