
レチノイドではなくビタミンAカロテノイドが、健康的な食事が腸内微生物の多様性に及ぼす影響を媒介する

リサーチ
公開日:2024年8月7日
レチノイドではなくビタミンAカロテノイドが、健康的な食事が腸内微生物の多様性に及ぼす影響を媒介する
BMC医学 22巻、記事番号:321(2024)この記事を引用する
131 アクセス
1 Altmetric
要旨
背景
ビタミンAは視力や免疫などの生理的プロセスに必須である。他のビタミンの吸収や代謝に影響を及ぼすビタミンAの腸内細菌叢組成への影響はまだ不明である。ここでは、腸内メタゲノム組成と6種類のビタミンA関連代謝産物(2種類のレチノイド:-レチノール、4種類のオキソレチノイン酸(oxoRA)、およびβ-クリプトキサンチンと3種類のカロテンジオールを含む4種類のカロテノイド代謝産物)の関係を調べた。
研究方法
TwinsUKコホートの1053人を対象とし、血清および糞便中のビタミンA関連代謝物、食歴、ショットガンメタゲノムシークエンシングによる腸内細菌叢組成を測定した。結果はZOE PREDICT-1研究の327人の女性で再現した。
結果
5種類のビタミンA関連血清代謝物がマイクロバイオームα多様性と正の相関を示した(r= 0.15 tor= 0.20,p< 4 ×10-6)。カロテノイド化合物は短鎖脂肪酸産生菌であるFaecalibacterium prausnitziiおよびCoprococcus eutactusと正の相関を示した。レチノールはどの微生物種とも関連していなかった。腸内細菌叢の組成は、ランダムフォレストモデルを用いて、カロテノイドとオキソレチノイン酸の循環レベルを0.66~0.74のAUCで予測することができたが、レチノール(AUC = 0.52)は予測できなかった。
健康的な食事指数(HEI)は、腸内細菌叢の多様性およびすべてのカロテノイド化合物と強く関連していたが、レチノイドは関連していなかった。健康的な食事(HEI)が腸内細菌叢の多様性に及ぼす影響に対するカロテノイド化合物の媒介的役割を調べたところ、カロテノイドはHEIが腸内細菌叢αの多様性に及ぼす影響の18~25%を有意に媒介することがわかった。
結論
我々の結果は、循環カロテン化合物と腸内細菌叢組成との間に強い関連性があり、健康的な食事パターンと関連する可能性があることを示している。
背景
ビタミンは、抗酸化作用や神経保護作用を有する微量栄養素である。これらの機能に加えて、一部のビタミンは腸内細菌叢の組成や多様性の変化と関連している [1] 。人体が適切な生理機能を果たすためには、13種類のビタミンが必要であり、それらは脂溶性ビタミン(ビタミンA、D、E、K)と水溶性ビタミン(ビタミンB群8種、ビタミンC)の2種類に分けられる。
脂溶性ビタミン、特にビタミンA、Dは、消化管で吸収され血流に分泌される際の免疫系への影響について研究されてきた [2]。567人の高齢者を対象とした最近の横断研究では、ビタミンD濃度が高い人ほど腸内の酪酸産生菌が豊富であることが判明した[3]。しかし、ビタミンAレベルと腸内細菌叢の関係については、まだ十分に研究されていない。
ビタミンAは、腸上皮バリアの完全性を維持するために不可欠な多量栄養素である [4,5]。ビタミンAは、粘液、タイトジャンクションタンパク質 [6]、抗菌ペプチドの産生と維持をサポートし、これら全てが病原性の侵入から腸内を守り、微生物叢の安定した環境を維持する。健康な腸管バリアは、ディスバイオシス(微生物の不均衡)を防ぎ、多様な微生物生態系を支えている [7]。さらに、ビタミンAには抗炎症作用があり [8]、腸内の慢性炎症を緩和するのに役立つ [9]。炎症を抑え、腸の上皮バリアをサポートすることで、ビタミンAはより安定した多様な微生物環境を育み、有益な常在菌を増やし、好ましくない常在菌を減らす。
ビタミンAは、動物性食品に含まれるレチニル酸、または植物性食品に含まれるカロテンやカロテノイドからの一連の酵素反応によって得られる。ビタミンAというとレチノールが連想され、実際、レチノールは人体におけるレチノイドの主要な形態であるが、主な生物学的活性分子は酸化誘導体である11-cis-レチナールと全トランスレチノイド酸(ATRA)である [10] 。ATRAは活性型として作用し、レチノイン酸受容体に結合する。
肝臓のビタミンAの80%以上は肝星細胞に貯蔵されている [11] 。肝細胞では、レチニルエステルヒドロラーゼによってレチニルエステルが加水分解され、レチノールが生成される。レチノールはその後、レチノール結合タンパク質(RBP)に結合してから循環中に放出され、STRA6などの膜受容体を介して全身の細胞に取り込まれる。レチノールの動員過程は、RBPの合成と分泌の速度を支配する変数によって厳密に制御されている [10] 。
ヒトの体内でビタミンAを生成することはできないため、動物性食品に含まれるプレフォームドビタミンAか、いくつかの果物や野菜に含まれるプロビタミンAカロテノイドとして、食事から摂取する必要がある [10]。牛乳や乳製品、肉類やその製品は、ヒトの食事に前形成ビタミンAを最も多く摂取する食品であり、次いで卵、卵製品、魚類である [10,13]。
カロテノイドは最終的にレチノールに代謝される [14] 。レチノイン酸は腸管免疫と透過性の重要な決定因子であることが示されている [15] 。重要なことは、食事性ビタミンAの吸収は胆汁酸の脂溶化特性 [16] に依存しており、胆汁酸は腸内細菌叢の組成や二次胆汁酸の細菌抱合 [17] によって調節されることである。
カロテノイドを多く含む食事は健康的な腸内細菌叢を維持する可能性が高く [18] 、ビタミンAは免疫機能と腸管バリアーの完全性に複数の有益な影響を及ぼす [18,19] ことから、α多様性とビタミンAには正の関係があると考えられた。
本研究の目的は、腸内細菌叢の組成やα多様性と、血流や便中のビタミンA関連代謝産物、特にレチノイドやカロテノイドの存在との間に関連があるかどうかを、機械学習法を用いて調査することである。さらに、胆汁酸がレチノイドとカロテノイドの濃度にどの程度影響を与えるかを定量化した。最後に、健康的食事指数(HEI)[20]と腸内細菌叢の構成との良好な関連において、ビタミンA代謝産物が果たす役割を調査する。
調査方法
発見コホート
研究参加者は、特定の疾患や形質を選択することなくボランティアとして募集された成人双子の全国登録であるTwinsUK登録に登録された個人である[21]。双子は文書によるインフォームドコンセントを提供し、本研究は聖トマス病院研究倫理委員会(REC Ref: EC04/015)によって承認された。ここでは、ビタミンA代謝物、質量分析による胆汁酸の測定、ショットガンメタゲノムによる腸内細菌叢のプロファイリングを同時に行った1053人を対象とした。対象者の一部は、HEI [20]によって測定された習慣的な食事の質に関するデータも入手可能であった。
再現コホート
再現コホートは、英国を拠点とするZOE PREDICT-1研究[22]の女性327人からなる独立したサンプルで構成され、ポストホック解析において、Metabolon Inc.による血清および便中ビタミンA代謝物の測定、ならびにホールショットガンメタゲノムデータが利用可能であり、食物摂取頻度調査票(FFQ)を記入した。ZOE PREDICT-1の倫理的承認は、St. Thomas Hospitalの研究倫理委員会から得た。全個人がインフォームド・コンセント(IRAS 236407)を行い、試験はClinicalTrials.govに登録された(登録番号:NCT03479866)。
ビタミンAメタボロミクスプロファイリング
ビタミンA濃度は、便および血清検体から、Metabolon Inc. 血清からは6種類のビタミンA関連代謝物が検出され、そのうち2種類のレチノールと4種類のカロテノイドが検出され、便からは5種類のビタミンA関連代謝物が検出された(レチノール1種類とカロテノイド4種類)。ビタミンAプロファイリングからバッチのばらつきを取り除くため、各ビタミンA関連代謝物について、実験サンプルの値を各装置バッチのサンプルの中央値で割り、各バッチ、したがって各ビタミンA代謝物の中央値を1とした。測定値が80%以上欠測したビタミンA代謝物は除外し、欠測率が20~80%のものは2分した。サンプルの80%以上に存在するビタミンA代謝物はバッチ正規化および逆正規化しました。洗浄後、血清中のビタミンA代謝物は6種類(レチノール2種類、カロテノイド4種類)、便中のビタミンA代謝物は5種類(レチノール1種類、カロテノイド4種類)残った。カロテノイド代謝物にはβ-クリプトキサンチン、カロテンジオール(1-3)が含まれる。レチノール代謝物には、4-オキソレチノール酸(oxoRA)とレチノールが含まれる。
胆汁酸メタボロミクスプロファイリング
コール酸(CA)、チェノデオキシコール酸(CDCA)、タウロコール酸(TCA)、グリココール酸(GCA)、タウロチェノデオキシコール酸(TCDCA)、グリコチェノデオキシコール酸(GCDCA)を含む循環一次胆汁酸は、上記のように、アンターゲットLC-MSプラットフォームを使用して、Metabolon Inc.
食事情報
European Prospective Investigations into Cancer and Nutrition(EPIC)-Norfolk研究 [25] のために確立された有効な131項目の半定量的食物摂取頻度調査票(FFQ)を用いて、習慣的な食事情報を推定した。FFQから、FETAソフトウェア[25,26]を用いて食品項目、マクロ栄養素および微量栄養素の摂取量を決定し、食品および栄養素の摂取量を特徴付け、様々な慢性疾患と関連することが理解されているHEI[27]を含む、食事パターン全体を表す指標を算出した。
マイクロバイオームの配列決定とプロファイリング
TwinsUKおよびZOE PREDICT-1の便サンプルのディープショットガンメタゲノム配列決定とそのプロファイリングは、以前に記載されたように実施した[28,29]。有病率の低い菌種によるノイズを減らし、統計的に有意な菌種を検出する力を向上させるために、有病率<20%の菌種を除去した。
統計解析
統計解析はStataバージョン18とRバージョン4.3.1を用いて行った[30]。
アルファ多様性については、個々の参加者内の腸内細菌叢の多様性を定量化するためのいくつかの指標を構築した。シャノンエントロピーとも呼ばれるシャノン多様性指標[31]は、エントロピーのクロード・シャノンの公式を用いて種の多様性を推定するものであり[22]、シンプソン多様性指標は、一般的な種や支配的な種に重みを与えるもので、Rパッケージ'vegan'の'diversity'関数を用いて計算した[32]。観察された種の単純な数である観察された豊かさは、「specnumber」関数を使って計算した。双生児であることを考慮し、年齢、性別、肥満度(BMI)を固定効果として、家族構成をランダム効果として調整した線形混合モデルを、TwinsUKとZOE PREDICT-1の両方で独立に用いて、ビタミンA関連代謝物と腸内細菌叢特性との単変量解析を行った。具体的には、いくつかのα多様性指標との関連を調べ、分類学的レベルでは逆正規化種存在量との関連を調べた。P値はBonferroni補正を用いて多重検定で補正し、補正後のP値が0.05未満の関連を有意とみなした。結果は固定効果メタアナリシスを用いて統合した。参加者間の差異(β多様性)とビタミンA関連代謝産物レベルを評価するため、腸内微生物群集データからBray-Curtis非類似度行列を構築し、年齢、性別、BMIを調整した上で、「adonis」関数('vegan'パッケージ[32])を用いたPERMANOVA(1000シミュレーション)を行い、有意性を判定した。
腸内メタゲノムデータを用いてビタミンA関連代謝物レベルをどの程度予測できるかを定量化するために、R [33]のrandomForestパッケージ(バージョン4.7-1.1)のランダムフォレスト(RF)モデルを用いた。モデルはTwinsUKとZOE-PREDICT-1のデータで独立に実施した。データセットをトレーニング・セット(80%)とテスト・セット(20%)に分割し,パフォーマンスをテストするために保持した.トレーニング・データでは,分散がゼロまたはゼロに近い予測変数は,'nearZeroVar'関数(caret R パッケージ[34])を使用して除外された.ハイパーパラメータ、mtry(各分割の候補としてランダムにサンプリングされた変数の数)、min.node.size(各ツリー中の終端ノードの最小サイズ)は、3回の繰り返しで、5倍適応リサンプリング(Rのcaretパッケージ[34])を使用してチューニングした。各モデルの最適な特徴数は、5重の再帰的特徴除去(randomForestパッケージ[33]の'rfcv'関数)によって決定された。モデルの性能をクロスバリデーションのエラー率で評価し、最適な特徴のサブセット(最小のエラー率で最小の特徴数)を決定することにより、繰り返し特徴を除去した。モデルをトレーニング・データで再トレーニングし、テスト・セットを使用して予測性能をテストした。各モデルの性能は、異なるクラスを識別するモデルの能力を測定する受信者動作特性下面積(AUC)と、モデルの予測クラスと真のクラスとの間のスピアマンの相関を用いて評価され、モデルの精度と信頼性が定量化された。
また、線形混合モデルを用いて、ビタミンA関連代謝物と(i)血清中の胆汁酸レベルおよび(ii)HEIとの関連を調べた。
ビタミンA関連代謝物が健康的な食事と腸内細菌叢の多様性との関係を媒介するかどうかを調べるために、Baron and Kennyアプローチ[35]を用いて因果媒介分析を行った。まず3つの媒介の仮定を検証し、次にRパッケージ'mediation'(バージョン4.5.0)の'mediate'関数を用いて因果媒介分析を行った[36]。各分析は各ビタミンA代謝物について独立に行い、年齢、性別、BMIで調整した。有意な媒介効果は、間接効果の有意性(p<0.05)と大きさによって決定された。さらに、間接効果と全体効果の比として、媒介過程によって説明される分散の割合、例えば、健康的な食事が腸内細菌叢に及ぼす影響のうち、ビタミンA代謝物を経由する割合などを表すVariance accounted for(VAF)を計算した。Vanderweeleの感度分析[37,38]を用いて、未測定の交絡に対する推定媒介効果の頑健性を評価した。これは、測定不能な交絡因子がメディエーター-アウトカム関係と曝露-メディエーター関係の両方に及ぼす潜在的な影響を評価するものである。
結果
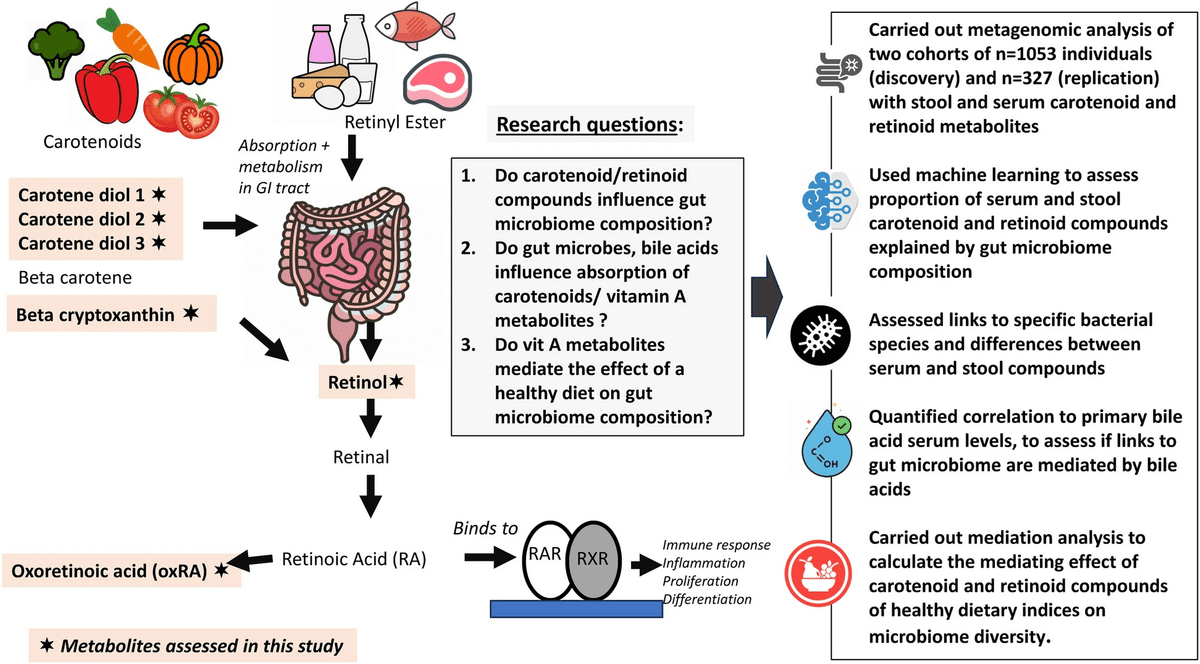
研究デザインのフローチャートを図1に示す。
図1
研究の概略図。この図では、食事摂取量と研究対象としたビタミンA関連代謝産物との関連を説明し、我々が取り組んでいる研究課題、データ、使用した分析ワークフローを強調している。
研究対象集団の記述的特徴を表1に示す。我々は、TwinsUKコホート[21]から1053人、ZOE PREDICT-1研究[21,22]から327人の女性を対象とし、Metabolon社[24]によって血清(n= 6)と便(n= 5)のビタミンA代謝物を測定した。[24]、腸内細菌叢組成はホールショットガンメタゲノミクスにより評価した。TwinsUKの平均(標準偏差、SD)年齢は57.72(15.20)歳、ZOE PREDICT-1の平均年齢は53.80(7.00)歳、平均体格指数(BMI)はそれぞれ26.18(5.15)kg/m2、26.24(5.62)kg/m2であった。
表1 研究集団の記述的特徴
ビタミンA代謝物と腸内マイクロバイオームとの関連
レチノールを除くすべてのビタミンA関連血清代謝物は、TwinsUKコホートにおいて年齢、性別、BMI、家族関係を調整した後、シャノン多様性指数と正の相関を示した(図2、パネルAおよびB)。さらに、心血管疾患(脳血管疾患、心不全、虚血性心疾患、冠動脈疾患、心房細動を含む)、2型糖尿病、慢性閉塞性肺疾患、アレルギー、食事(健康的な食事指数、食物繊維摂取量、野菜摂取量、エネルギー摂取量を含む)、抗生物質の使用、身体運動、ビタミン補給、塩基配列の深さについて調整しても、結果は変わらなかった(追加ファイル1:TableS1)。ZOE-PREDICT 1(図2、パネルAおよびB)およびシンプソン指数および観察された種の数との相関を調査した場合、一貫した結果が観察された(追加ファイル1:表S2)。さらに、年齢、性別、BMIで調整した後、ビタミンA代謝物とβ多様性(Bray-Curtis指標で決定)との間に有意な関係が検出された(Additional File 1:TableS3)。
図2
血清および便からのビタミンA代謝物とシャノン多様性指数(AおよびB)および健康的食事指数(CおよびD)との関連。データを示す。丸印はTwinsUKコホートとZOE PREDICT 1コホートにおける効果量、菱形は両コホートを組み合わせた固定効果メタ解析(MA)から得られた要約効果量。ひげは95%信頼区間を表す。統計的有意性は以下のようにアスタリスクで示されている: *p< 0.05; **p< 0.01; ***p< 0.001; ****p< 0.0001
次に、TwinsUKにおいて、ビタミンA関連代謝物と有病率20%以上の腸内細菌種との単変量解析を行ったところ、49の固有の細菌種を含む96の有意な関連(Bonferroni < 0.05)が同定された(図3A、追加ファイル1:表S4)。便の代謝物レベルとの関連も同様であったが弱かった(図3A)。関連する菌種のうち、Faecalibacterium prausnitziiと Coprococcus eutactusはすべてのカロテノイド代謝物と正の相関を示したが、負の相関を示したのはRuminococcus torquesや R. gnavus、Eggerthella lenta、 Tyzzerella nexilis、Flavonifractor plautiiなどであった。個々の分類群と血清または便中のレチノール濃度との間には関連は認められなかった。
図3
血清と腸内細菌種から得られたビタミンA関連代謝物の相関を表したヒートマップ。マトリックスの各セルには、ビタミンA関連代謝物と細菌種との回帰係数が示されている。表は凡例に従って相関ごとに色分けされている(赤は正の相関、青は負の相関)。*p< 0.05; **p< 0.01; ***p< 0.001; ****p< 0.0001。B TwinsUKおよびZOE PREDICT-1参加者における、ランダムフォレスト回帰器(スピアマンの相関を使用)および分類器(AUC値を使用)によって推定されたビタミンA関連代謝産物における腸内細菌叢の予測。箱ひげ図は、TwinsUKとZOE PREDICT-1の平均AUCと5倍にわたる95%信頼区間を表す。紺色と水色の円は、TwinsUKとZOE PREDICT-1における、各ビタミンA関連代謝物の実際の値と、5倍にわたるモデルによる予測値との間のスピアマンの相関の平均を示す。
次に、ランダムフォレストモデルを用いて、微生物種の存在量が循環ビタミンA関連代謝物レベルを予測する能力を測定し、腸内細菌叢組成がビタミンA代謝物レベルとどの程度関連しているかを定量的に推定した。モデルの性能は、受信者動作特性曲線下面積(AUC)を用いて評価し、予測値と観察値の間のスピアマンの相関(Rho)を算出した。TwinsUKでは、平均して、腸内細菌叢は5フォールドにわたってカロテノイド代謝物の循環レベルを予測することができ(図3B)、カロテンジオール(2)が最も強い関連を示した(AUC[95%CI]=0.74[0.67;0.81]、rho[95%CI]=0.31[0.15;0.45])。
感度分析を行い、結果が腸内細菌叢の変換(逆正規化)に依存するかどうかを調べた。中心対数比(CLR)変換を用いてデータを変換したところ、結果は一貫していた(例えば、カロテンジオール(2)のAUC[95%CI]=0.68[0.6;0.76]、rho[95%CI]=0.25[0.1;0.4])。
予測モデルをZOEPREDICT-1コホートで再現した。TwinsUKでの観察結果と一致して、腸内細菌叢はカロテノイド代謝物の循環レベルを予測することができ、カロテンジオール(2)のAUCが最大であった(0.73 [0.60, 0.86])。
ビタミンA代謝物と胆汁酸レベル
TwinsUKでは、一次胆汁酸によって調節される脂肪吸収が血清ビタミンA濃度に影響を及ぼすかどうかを調べるために、カロテノイドとレチノイドの循環量に対する一次胆汁酸の影響を定量化した[4]。CA、CDCA、およびそれらの共役胆汁酸塩であるTCA、GCA、TCDCA、GCDCAとの相関を検証した。これらの胆汁酸塩は、ビタミンAの吸収と代謝に影響を及ぼすことが文献で示唆されており [16]、腸内細菌叢の構成と機能に強い関連があることが知られている [39,40]。カロテンジオール(2)はGA(β[95%CI]=-0.08[-0.14、-0.07]、p=0.01)およびGCDCA(β[95%CI]=-0.06[-0.12、-0.004]、p=0.03)と負の相関があった。 03)、カロテンジオール(1)はCAと正の相関を示し(β[95%CI]=0.07[0.001,0.15]、p=0.05)、oxoRAはCAと正の相関を示した(β[95%CI]=0.07、[0.001,0.16]、p=0.045、それぞれ0.12[0.05,0.12]、p=5.43×10-4、追加ファイル1:図1)。多重検定(Bonferronip値=0.05/30=1.6×10-3)で調整した結果、oxoRAのみがCAレベルと関連していた。どのカロテノイドもレチノールも、多重検定で調整した後では一次胆汁酸と相関しなかった。
ビタミンA代謝物と食事の質との関連
さらに、血清および便中ビタミンA関連代謝物と、HEIで測定した習慣的な食事の質との関連を、FFQが利用可能なTwinsUKの664人のサブセットで調べた。共変量で調整した後、血清カロテノイドとHEIとの間に強い正の相関が認められたが、レチノールや便中ビタミンA代謝物量には影響は認められなかった(図2C)。結果はPREDICT 1でも再現された(図2D)。
文献[41]と一致して、年齢、性別、BMI、家族関係を調整した後、HEIとシャノン多様性指数(β [95% CI] = 0.10 [0.02, 0.19]、 p= 0.01)の間に正の関連が認められた。重要なことは、各カロテノイドを順に調整すると、その効果は減弱したことである(追加ファイル1:表S5)。そこで、年齢、BMI、性別を調整した正式な媒介分析を行い、血清カロテノイド代謝物がHEIとシャノン多様性指数との関連を媒介するかどうかを調べた。嫌気的条件下では(ヒトの大腸のように)腸内微生物はカロテノイドを産生できないことが実験的に証明されているためである[42,43])。4つのカロテノイド代謝物はすべて、BaronとKenny [35]が示したメディエーターの基準を満たしていた。正式な媒介分析によると、4つのカロテノイドはすべて、腸内微生物の多様性に対する健康的な食事(HEI)の効果を有意に媒介した(図4)。説明された分散の範囲は、β-クリプトキサンチンの19.8%からカロテンジオールの39.9%であった(1)。
図4
媒介分析図。健康的な食事は、健康的な食事指数で表され、曝露としてモデル化された。シャノン多様性指数で測定した腸内細菌叢のα多様性をアウトカムとしてモデル化した。各ビタミンA代謝物をメディエーターとしてモデル化した。媒介分析の要約統計量はベータ値(95%信頼区間)として描かれている。略語 HEI、健康的な食事指数、CI、信頼区間
しかし、カロテンジオール(1)と(2)を取り巻く我々の知見は、測定されていない残留交絡の影響を受けやすい可能性がある(e値=1.32 [LowerBound = 1.0] [37,38] (Additional File 1:TableS6))。一方、カロテンジオール(3)とβ-クリプトキサンチンについては、結果は頑健であった(e-values= 1.35 [LowerBound = 1.04])。
考察
便および血清中のビタミンA関連代謝産物と腸内細菌叢との関連を調べたこの大規模研究において、カロテノイドは腸内細菌叢の構成と強い相関がある一方、レチノイドは腸内細菌と弱い相関しかないことが報告された。
アルファ多様性との強い正の相関に加えて、カロテノイド化合物の循環レベルと、Faecalibacterium prausnitziiや Coprococcus eutactusなどの短鎖脂肪酸(SCFA)産生腸内微生物の存在量との間にも有意な正の相関が観察された[44,45,46]。カロテノイドには抗酸化作用があることが知られており [47]、有益な細菌種がより多く存在することに寄与している可能性がある。同様に、カロテノイドは、大腸がんのリスクや進行(Eggerthella lenta、 Tyzzerella nexilis、 Flavonifractor plautii)[49,50,51]だけでなく、以前から炎症(Ruminococcus torques、Ruminococcus gnavus)[48]や好ましくない心代謝系の結果[29]に関連している菌種と負の相関関係があることがわかった。
一貫して、血清中のこれらの化合物のレベルは、便中でははるかに少ないが、ランダムフォレストモデルを使用して腸内細菌叢の組成によって部分的に予測され、重要なことに、これらの結果は独立したコホートで再現された。カロテノイドとレチノイドの便中濃度に関する腸内細菌叢の予測が欠如していることは、これらの物質の吸収が主に近位腸(空腸と十二指腸)で行われることを反映している可能性がある [52] 。
また、これらの化合物はすべて脂溶性であるが、一次胆汁酸(脂肪物質の吸収において重要であり、ビタミンAとその代謝物の吸収・代謝に関与している [16])のレベルは、カロテノイド化合物やレチノールの循環レベルと相関していないことも報告した。胆汁酸代謝と腸内細菌叢の構成との間には強い関連性が報告されているが [17,40]、今回報告された腸内細菌叢の多様性との関連は、上部消化管での吸収に対する腸内細菌叢の影響によるものとは考えにくく、このメカニズムは、健康な集団でみられる範囲内の循環カロテノイドおよびレチノイドのレベルを決定する上で、わずかな役割しか果たしていない可能性を示唆している。実際、最近の実験データから、腸内微生物はヒトの腸に匹敵する嫌気的条件下ではカロテノイドを合成できないことが示されており[43]、したがって、我々のデータは、食事中のカロテノイドが腸内細菌叢の構成に影響を与えるのであって、その逆ではないことと一致している。
最後に、健康的な食事(HEIの遵守によって測定)が腸内細菌叢のα多様性に及ぼす影響は、ある程度カロテノイド化合物のレベルによって媒介されることが示された。その割合は、β-クリプトキサンチンの19.8%からカロテンジオールの39.9%であった(1)。レチノイド化合物には有意な効果は見られなかった。これらの結果は、個別化栄養学や、健康な腸内細菌叢を促進するための食事介入法の開発に潜在的な意味を持つ可能性がある [53] 。実際、これらのデータは、脂溶性カロテノイドと有益な腸内細菌との間に強い関連があることを強調しており、カロテノイド摂取によってこれらの種の一部を調節できる可能性を示唆している。この分野の研究がさらに進めば、腸内微生物の組成を最適化し、全体的な健康を促進することを目的とした、個人に的を絞った推奨食の開発に貢献できるかもしれない。
我々の研究には多くの長所がある。第一に、十分に検証された大規模集団ベースコホートを探索に用い、独立したコホートで得られた知見を検証したことである。両コホートとも同じオミックスプロファイリングを行い、同じ品質管理と解析パイプラインを用いて処理した。第二に、16S rRNA遺伝子のアンプリコンデータを用いた解析よりもはるかに深度の深い全ショットガンメタゲノミクスデータを用いた。第三に、市販されているメタボロミクスパネルの中で最も優れた特性を持つ標的メタボロミクスパネルの1つを用いて、血清と便の両方でビタミンA代謝物と数種類の一次および二次胆汁酸を測定した。
また、いくつかの制限もある。第一に、本研究のサンプルは女性が多く、再現コホートは女性のみであった。第二に、使用したメタボロミクスパネルが提供するビタミンA代謝物濃度は相対値であり、血清中または便中の代謝物濃度の絶対値ではない。この限界は、ビタミンA欠乏症やビタミンA中毒の診断/管理に用いられる臨床的閾値と比較するために絶対定量が必要な、我々の結果の臨床的解釈可能性にも影響する。とはいえ、われわれの結果は相対的な変化と関連性を示しており、今後の研究でより深く検討し、臨床的妥当性を向上させることが可能である。第3に、食事摂取量は食物摂取頻度調査票を用いて測定されたが、これには回答バイアスや誤分類の可能性を含む固有のバイアスがある。第4に、我々のメディエーション解析では、曝露、メディエーター、アウトカムの測定間に時間的関係がないため、因果関係を真に同定できない可能性があり、測定されていない交絡因子が残存している可能性を排除できない。それにもかかわらず、我々はTwinsUKとZOE Predict-1の両方で結果を再現することに成功した。最後に、カロテノイドレベル、腸内細菌叢組成、HEI間の因果関係を推論することはできなかった。
結論
まとめると、これは我々の知る限り、全ゲノムショットガンメタゲノムシーケンスを用いてビタミンA代謝物と腸内細菌叢の関連を調査した初めての大規模研究である。ここで我々は、カロテノイドはレチノイドではなく、より高いアルファ多様性およびより高い善玉菌相対量と強い相関があること、そしてビタミンA代謝産物が、健康的な食生活の遵守がマイクロバイオームの多様性に及ぼす影響の20~39%を媒介することを示した。
データおよび資料の入手可能性
本研究で使用したデータは、キングス・カレッジ・ロンドンのDepartment of Twin Researchが保有している。データは、Wellcome Trustとそのガイドラインによって監督された通常の手順を用いて、私たちのコアファンディングの一環として善意の研究者に公開することができます(https://twinsuk.ac.uk/resources-for-researchers/access-our-data/)。腸内細菌叢のデータは、EBI(https://www.ebi.ac.uk/)のアクセッション番号PRJEB39223(ZOE- PREDICT-1)およびPRJEB32731(TwinsUK)で入手可能である。
略号
ATRA:
オールトランスレチノイド酸
BMI:
体格指数
CA:
コール酸
CDCA
チェノデオキシコール酸
FFQ:
食物摂取頻度調査票
GCA
グリココール酸
GCDCA
グリコヘノデオキシコール酸
HEI
健康的な食事指数
oxoRA
4-オキソレチノール酸
RAR
レチノイン酸受容体
RBP
レチノール結合タンパク質
TCA
タウロコール酸
TCDCA
タウロヘノデオキシコール酸
参考文献
Rinninella E, Mele MC, Merendino N, Cintoni M, Anselmi G, Caporossi A, et al. 加齢黄斑変性における食事、微量栄養素、腸内細菌叢の役割: 腸-網膜軸からの新たな展望。Nutrients. 2018;10:1677.
Mora JR, Iwata M, von Andrian UH. ビタミンの免疫系への影響:ビタミンAとDが主役。Nat Rev Immunol. 2008;8:685-98.Article CAS PubMed PubMed Central Google Scholar
高齢男性におけるビタミンD代謝産物と腸内細菌叢。Nat Commun. 2020;11:5997.Article CAS PubMed PubMed Central Google Scholar
代謝調節における胆汁酸の役割。J Endocrinol。2016;228:R85-96.Article PubMed Google Scholar
ビタミンAとビタミンDは、微生物の複雑性、バリア機能、粘膜免疫応答を制御し、腸の恒常性を確保する。Crit Rev Biochem Mol Biol. 2019;54:184-92.Article CAS PubMed PubMed Central Google Scholar
Lima AAM, Soares AM, Lima NL, Mota RMS, Maciel BLL, Kvalsund MP, et al. ブラジルの小児における腸管バリア機能、成長、総寄生虫、特異的ジアルジア属感染に対するビタミンA補給の効果:前向き無作為化二重盲検プラセボ対照試験。J Pediatr Gastroenterol Nutr. 2010;50:309-15.Article CAS PubMed PubMed Central Google Scholar
Stolfi C、Maresca C、Monteleone G、Laudisi F.腸内バリア機能不全の腸内dysbiosisおよび疾患における含意。Biomedicines. 2022;10:10.Article Google Scholar
抗炎症剤としてのビタミンA。論文 CAS PubMed Google Scholar
慢性炎症性疾患における宿主免疫細胞と腸内細菌叢の相互作用。Exp Mol Med. 2017;49: e339.論文 CAS PubMed PubMed Central Google Scholar
ビタミンAの最新情報:形態、供給源、動態、検出、機能、欠乏、治療使用および毒性。栄養素。2021;13:1703.Article CAS PubMed PubMed Central Google Scholar
ビタミンAの吸収、貯蔵および動員。ビタミンAの吸収、貯蔵、動員。Subcell Biochem. 2016;81:95-125.Article CAS PubMed Google Scholar
Bawa FNC, Zhang Y. 脂肪性肝疾患におけるレチノイン酸シグナル。Liver Res. 2023;7:189-95.Article PubMed PubMed Central Google Scholar
アルビナリHAH。夜盲症と古代の治療法。ハートビュー。2014;15:136-9.Article PubMed PubMed Central Google Scholar
脂肪細胞機能、脂肪率と肥満の制御におけるカロテノイドとその変換産物。Arch Biochem Biophys. 2015;572:112-25.Article PubMed Google Scholar
Bakdash G, Vogelpoel LTC, van Capel TMM, Kapsenberg ML, de Jong EC. レチノイン酸はヒト樹状細胞をプライミングし、腸管ホーミング性IL-10産生制御性T細胞を誘導する。Mucosal Immunol. 2015;8:265-78.Article CAS PubMed Google Scholar
Saeed A, Hoekstra M, Hoeke MO, Heegsma J, Faber KN. 胆汁酸とビタミンAの恒常性の相互関係。Biochim Biophys Acta Mol Cell Biol Lipids. 2017;1862:496-512.Article CAS PubMed Google Scholar
Staley C, Weingarden AR, Khoruts A, Sadowsky MJ. 腸内細菌叢と胆汁酸代謝との相互作用と病態への影響。Appl Microbiol Biotechnol. 2017;101:47-64.Article CAS PubMed Google Scholar
Eroglu A, Al'Abri IS, Kopec RE, Crook N, Bohn T: Carotenoids and Their Health Benefits as Derived via Their Interactions with Gut Microbiota. 2023;14(2):238-55.
McCullough FS, Northrop-Clewes CA, Thurnham DI. 上皮の完全性に対するビタミンAの効果。Proc Nutr Soc. 1999;58:289-93.Article CAS PubMed Google Scholar
Guenther PM, Casavale KO, Reedy J, Kirkpatrick SI, Hiza HAB, Kuczynski KJ, et al. 健康的な食事指数の更新: HEI-2010。J Acad Nutr Diet. 2013;113:569-80.Article PubMed Google Scholar
TwinsUK: the UK adult twin registry update. Twin Res Hum Genet. 2019;22:523-9.Article PubMed Google Scholar
Berry SE, Valdes AM, Drew DA, Asnicar F, Mazidi M, Wolf J, et al. ヒトの食後反応と精密栄養学の可能性。Nat Med. 2020;26:964-73.Article CAS PubMed PubMed Central Google Scholar
Nogal A, Tettamanzi F, Dong Q, Louca P, Visconti A, Christiansen C, et al.空腹時血糖障害の糞便代謝物シグネチャー:2つの独立した集団ベースのコホートからの結果。糖尿病。2023;72:1870-80.Article CAS PubMed PubMed Central Google Scholar
糖尿病と慢性腎臓病の関連性を検討した。2024;27(3):109132.
Bingham SA, Welch AA, McTaggart A, Mulligan AA, Runswick SA, Luben R, et al. Norfolkにおけるがんの欧州前向き調査における栄養学的方法。Public Health Nutr. 2001;4:847-58.Article CAS PubMed Google Scholar
Mulligan AA, Luben RN, Bhaniani A, Parry-Smith DJ, O'Connor L, Khawaja AP, et al. 食品摂取頻度調査票データを栄養素および食品群値に変換するための新しいツール: FETAの調査方法と利用可能性。BMJ Open. 2014;4: e004503.記事 PubMed PubMed Central Google Scholar
ケネディET、オールズJ、カールソンS、フレミングK.健康的な食事指数:デザインとアプリケーション。J Am Diet Assoc. 1995;95:1103-8.Article CAS PubMed Google Scholar
ヒトの腸内細菌叢と宿主の代謝との相互作用。Nat Commun. 2019;10:4505.https://doi.org/10.1038/s41467-019-12476-z.Article CAS PubMed PubMed Central Google Scholar
Asnicar F, Berry SE, Valdes AM, Nguyen LH, Piccinno G, Drew DA, et al. 1,098人の深い表現型から得られたマイクロバイオームと宿主代謝および習慣的食事との関連。Nat Med. https://doi.org/10.1038/s41591-020-01183-8.Article CAS PubMed PubMed Central Google Scholar
統計計算のためのRプロジェクトhttps://www.R-project.org. 2024年1月30日アクセス。
Konopiński MK. Shannon diversity index: a call to replace the original Shannon's formula with unbiased estimator in the population genetics studies. PeerJ. 2020;8: e9391.記事 PubMed PubMed Central Google Scholar
Community Ecology Package [R package vegan version 2.6-6.1]. 2024.
遼A. randomForestによる分類と回帰. R news. 2002;2(3):18-22.Google Scholar
Kuhn M. Building predictive models in R using the caret package. J stat softw. 2008;28:1-26.Article Google Scholar
Baron RM, Kenny DA. 社会心理学研究におけるモデレーター変数とメディエーター変数の区別:概念的、戦略的、統計的考察。J Pers Soc Psychol. 1986;51:1173-82.Article CAS PubMed Google Scholar
Tingley D, Yamamoto T, Hirose K, Keele L, Imai K. Mediation: 因果媒介分析のためのRパッケージ。J Stat Softw. 2014;59:1-38.Article Google Scholar
Smith LH, VanderWeele TJ. Mediational E-values: approximate sensitivity analysis for unmeasured mediator-outcome confounding. Epidemiology. 2019;30:835-7.Article PubMed PubMed Central Google Scholar
VanderWeele TJ. 直接効果と間接効果の感度分析のためのバイアス公式。Epidemiology. 2010;21:540-51.Article PubMed PubMed Central Google Scholar
桐山泰弘、野地秀樹。腸内細菌叢によって修飾される胆汁酸の生理的役割。微生物.2021;10:10.Article Google Scholar
胆汁酸の生理的役割は、腸内細菌叢によって改変される。Cell Rep Med. 2023;4(4):100993.
Bowyer RCE, Jackson MA, Pallister T, Skinner J, Spector TD, Welch AA, et al. ヒト腸内細菌叢研究における食事のコントロールのための食事指標の使用。Microbiome. 2018;6:77.Article PubMed PubMed Central Google Scholar
Lin D, Medeiros DM. 消化管の主要機能としてのマイクロバイオームと微量栄養素代謝および慢性疾患への関与。Nutr Res. 2023;112:30-45.Article CAS PubMed Google Scholar
ヒト腸内細菌Eubacterium limosumおよびLeuconostoc mesenteroidesにおけるカロテノイド生産性とカロテノイド生合成遺伝子の機能解析. 工学微生物学。2024;4: 100147.Article CAS Google Scholar
ヒトの健康への応用が期待される細菌属、ファエカリバクテリウム(Faecalibacterium)。FEMS Microbiol Rev. 2023;47(4):fuad039.Article PubMed PubMed Central Google Scholar
腸内細菌が内臓脂肪に及ぼす影響を短鎖脂肪酸アセテートの循環レベルが仲介している。Front Microbiol. 2021;12: 711359.Article PubMed PubMed Central Google Scholar
潰瘍性大腸炎患者における酪酸産生菌Roseburia hominisとFaecalibacterium prausnitziiの減少が腸内細菌叢の異常を規定する。Gut. 2014;63:1275-83.Article CAS PubMed Google Scholar
Young AJ, Lowe GL. カロテノイドの抗酸化作用。抗酸化物質(バーゼル)。2018;7:7.Google Scholar
Hall AB, Yassour M, Sauk J, Garner A, Jiang X, Arthur T, et al. 炎症性腸疾患患者に濃縮された新規Ruminococcus gnavusクレード。Genome Med. 2017;9:103.Article PubMed PubMed Central Google Scholar
Bai X, Wei H, Liu W, Coker OO, Gou H, Liu C, et al. タバコの煙は腸内細菌叢と関連代謝産物の調節を介して大腸がんを促進する。Gut. 2022;71:2439-50.Article CAS PubMed Google Scholar
Lozano CP, Wilkens LR, Shvetsov YB, Maskarinec G, Park S-Y, Shepherd JA, et al. 多民族コホート脂肪率表現型研究における腸内細菌叢、LPS、C反応性タンパク質を介した食事性炎症指数と総脂肪率および異所性脂肪との関連。Am J Clin Nutr. 2022;115:1344-56.Article CAS PubMed Google Scholar
ALSを制御する腸内細菌叢と代謝産物の役割の可能性(Blacher E, Bashiardes S, Shapiro H, Rothschild D, Mor U, Dori-Bachash M, et al. Nature. 2019;572:474-80.Article CAS PubMed Google Scholar
Goncalves A, Roi S, Nowicki M, Dhaussy A, Huertas A, Amiot M-J, et al. 脂溶性ビタミンの腸管吸収:腸内の吸収部位と吸収の相互作用. Food Chem. 2015;172:155-60.Article CAS PubMed Google Scholar
ゴーシュTS、バルデスAM. 心代謝性疾患における腸内細菌叢を標的とした臨床介入のエビデンス。BMJ. 2023;383: e075180.Article PubMed PubMed Central Google Scholar
謝辞
TwinsUKおよびZOE-PREDICT-1に参加されたすべての方々(食事介入試験に時間を割いてくださった方々を含む)に心から感謝いたします。また、各コホートに従事したスタッフ、Zoe Global Limitedの研究運営とデータ収集に貢献した精力的な活動に感謝する。
資金提供
本研究の全部または一部はWellcome Trust(WT212904/Z/18/Z)の助成を受けた。オープンアクセスを目的として、著者らはこの投稿から生じたAuthor Accepted ManuscriptのバージョンにCC BYのパブリック著作権を適用した。双子研究部門は、Wellcome Trust (212904/Z/18/Z)、Medical Research Council (MRC)/British Heart Foundation (BHF)のAncestry and Biological Informative Markers for Stratification of Hypertension (AIM-HY; MR/M016560/1)、欧州連合、慢性疾患研究基金(CDRF)、Zoe Global Ltd.、NIHR Clinical Research Facility (NIHR臨床研究施設)からの助成を受けている、 NIHR Clinical Research Facility and Biomedical Research Centre(Guy's and St Thomas' NHS Foundation Trustを拠点とし、King's College Londonと提携)。CMはChronic Disease Research Foundation(CDRF)およびMRC Aim-Hyプロジェクト助成金、AMVはNational Institute for Health and Care Research Nottingham Biomedical Research Centreの助成を受けている。PLはCDRFの助成を受けている(CDRF-15/2018)。本研究は、AMVとCMへのUKRI/MRC助成金MR/W026813/1とMR/Y010175/1からも支援を受けた。
著者情報
著者情報
Ana M. ValdesとPanayiotis Loucaは本研究に等しく貢献した。
著者および所属
ノッティンガム医学部NIHR生物医学研究センター、ノッティンガム大学、ノッティンガム、NG5 1PB, UKAna M. Valdes
ノッティンガム大学医学部炎症・回復・傷害科学部門(英国、ノッティンガム、NG7 2UH)Ana M. Valdes
双生児研究・遺伝疫学部門、キングス・カレッジ・ロンドン、ロンドン、SE1 7EH、UKAna M. Valdes、Panayiotis Louca、Alessia Visconti、Ana Nogal、Tim D. Spector、Mario Falchi、Cristina Menni
人間栄養・運動研究センター、ニューカッスル大学、ニューカッスル・アポン・タイン、NE2 4HH, UKPanayiotis Louca
トリノ大学臨床・生物科学部生物統計・疫学・公衆衛生センター(イタリア・トリノ、10124)Alessia Visconti
トレント大学CIBIO学部、Via Sommarive 9、38123、Povo、Trento、ItalyFrancesco Asnicar & Nicola Segata
栄養科学科、キングス・カレッジ・ロンドン、150 Stamford Street, London, SE1 9NH, UKケイト・バーミンガム&サラ・E・ベリー
Zoe Limited, 164 Westminster Bridge Rd, London, SE1 7RW, UKケイト・バーミンガム、ジョナサン・ウルフ、ティム・D・スペクター、サラ・E・ベリー
ハーバードT.H.チャン公衆衛生大学院疫学科(マサチューセッツ州ボストン、02115、USA)アナ・ノガル(Anna Nogal
Metabolon Inc, Research Triangle Park, Morrisville, NC, 27560, USAKari Wong & Gregory A. Michelotti
貢献
実験の構想および設計: AMVとCM。試薬、材料、解析ツールの提供: AV、FA、KB、AN、KW、GAM、JW、NS、TDS、SEB、MF。データ管理: AV、FA、PL、CM。形式分析: PL、CM。原稿執筆: AMVとCM。原稿の修正:全員。最終原稿は著者全員が読み、承認した。
対応する著者
Ana M. ValdesまたはCristina Menniまで。
倫理宣言
倫理承認と参加同意
参加者全員が書面によるインフォームドコンセントを行った。TwinsUKはSt Thomas' Hospital Research Ethics Committeeの承認を得た(REC Ref: EC04/015)。ZOE PREDICT-1の倫理的承認はセント・トーマス病院研究倫理委員会から得られ、全個人が書面によるインフォームド・コンセントを行い(IRAS 236407)、試験はClinicalTrials.govに登録された(登録番号:NCT03479866)。
出版に関する同意
該当なし。
競合利益
TDSはZOE社の共同設立者であり株主である。AMV、SEB、KB、FA、NSはゾーイ社(以下「ゾーイ社」)のコンサルタントである。JWはゾーイ社の従業員。KWとGAMはMetabolon Inc.の従業員である。その他の著者は、競合する利害関係がないことを宣言している。
補足情報
追加情報
出版社ノート
シュプリンガー・ネイチャーは、出版された地図の管轄権の主張および所属機関に関して中立を保っています。
補足情報
12916_2024_3543_MOESM1_ESM.docx
補足資料1:表S1. 年齢、性別、BMI、心血管疾患(すなわち、脳血管疾患、心不全、虚血性心疾患、冠動脈疾患、心房細動)、2型糖尿病、慢性閉塞性肺疾患、アレルギー、食事(すなわち、健康的な食事指数、繊維摂取量、野菜摂取量、エネルギー摂取量)、抗生物質の使用、身体運動、ビタミンの補充、配列の深さ、TwinsUKにおける家族関連性を調整したシャノン多様性とビタミンA関連代謝産物との関連。この分析では、欠測した食事値は平均値にインプットされた。表S2. TwinsUKおよびPREDICT-1における、年齢、性別、BMI、家族関係を調整したα多様性指標とビタミンA関連代謝産物との関連。固定効果メタ解析のベータ(SE)とP値を報告。表S3. PERMANOVA(Bray-Curtis非類似度)を用いた血清および便中ビタミンA関連代謝産物との関連における腸内細菌叢組成のβ多様性推定値。ADONISに実装されているPERMANOVA解析の結果で、Bray-Curtis非類似度を用いた腸内細菌叢のβ多様性と血清および便のビタミンA関連代謝物との関連を調べた。表には、共変量(年齢、性別、BMI、家族関係)で未調整および調整した各代謝物のR2乗(R²)値およびp値が含まれる。R²値は、各代謝物によって説明される腸内細菌叢組成の全変動の割合を示す。P値は1000回の並べ替えから導き出され、観察された関連性の統計的有意性を示す。表S4. TwinsUKにおける、年齢、性別、BMI、家族関係、多重検定で調整した循環ビタミンA関連代謝物と細菌種との関連。表S5. 年齢、性、BMI、家族関係、以下のカロテノイドで調整したシャノンとHEIとの関連。表S6. E値法による測定不能交絡の感度分析。図S1. ビタミンA関連代謝物と循環一次胆汁酸との関連。
権利と許可
オープンアクセス本論文は、クリエイティブ・コモンズ表示4.0国際ライセンスの下でライセンスされている。このライセンスは、原著者および出典に適切なクレジットを与え、クリエイティブ・コモンズ・ライセンスへのリンクを提供し、変更が加えられた場合を示す限り、いかなる媒体または形式においても使用、共有、翻案、配布、複製を許可するものである。この記事に掲載されている画像やその他の第三者の素材は、その素材へのクレジット表記に別段の記載がない限り、記事のクリエイティブ・コモンズ・ライセンスに含まれています。この記事のクリエイティブ・コモンズ・ライセンスに含まれていない素材で、あなたの意図する利用が法的規制によって許可されていない場合、あるいは許可された利用を超える場合は、著作権者から直接許可を得る必要があります。このライセンスのコピーを閲覧するには、http://creativecommons.org/licenses/by/4.0/。クリエイティブ・コモンズ・パブリック・ドメインの権利放棄(http://creativecommons.org/publicdomain/zero/1.0/)は、データへのクレジット表記に別段の記載がない限り、この記事で利用可能となったデータに適用される。
この記事について
この記事の引用
Valdes,A.M.、Louca,P.、Visconti,A.et al.ビタミンAカロテノイドは、レチノイドではなく、健康的な食事が腸内微生物の多様性に与える影響を媒介する。BMC Med 22, 321 (2024). https://doi.org/10.1186/s12916-024-03543-4
2024年4月10日受領
受理2024年7月28日
2024年8月07日発行
DOIhttps://doi.org/10.1186/s12916-024-03543-4
この記事を共有する
以下のリンクを共有した人は、このコンテンツを読むことができます:
提供:Springer Nature SharedIt コンテンツ共有イニシアチブ
キーワード
BMC医学
ISBN: 1741-7015
お問い合わせ
投稿に関するお問い合わせ:bmcmedicineeditorial@biomedcentral.com
一般的なお問い合わせ:info@biomedcentral.com
BMCをフォローする
このウェブサイトを使用することで、当社の利用規約、お客様の米国州プライバシー権、プライバシーステートメント、およびクッキーポリシーに同意したものとみなされます。プライバシーに関する選択/プリファレンスセンターで使用するクッキーの管理。
特に明記されていない限り、© 2024 BioMed Central Ltd. シュプリンガー・ネイチャーの一部です。