
SIZE〜HYDRA

point
①スプライジング機能により遺伝子的超極所的な変化は断続的に進んでいる。
②生命の多様性=細胞の可能性指数の最大化=プラスミド=全能的母胎概念。
③減数分裂は初めからその形で存在したのではなく、個別機能の集合サイクル。
④幹細胞も、全能性細胞も初めから存在したわけではなく、後続的に出現した細胞。
⑤生殖系染色体は癌抑制遺伝子を持つ通常染色体がその進化種の中で不要となった
染色体から最低限の情報を残して縮合される。
⑥生命種は遺伝子の水平移動により多様化し、外的環境の条件により
効率的肥大化を目的とし、結果的にそれはよりコンパクトなものが選択される。
*二杯葉目が単離した時代、まだ脳は生成されていなかったが、その後明らかに
脳が作られる胚葉により睡眠という生体周期を身につけた。
これは意識が生まれた起源かもしれない。
睡眠の意味と理由、それは意識の精査を生み出す機能前置き
生命は偶発的な理由で進化したりしない、
突然変異は進化などではなく、あくまでも、エネルギー的秩序のもと産み落とされる一つの生命種に過ぎない。
進化とはほぼ明確に意思を持つ、
その意思の源は
空間や、時間、などといった物理的かつ感覚的概念の機能的発現とともに発生し、組織の増大化と染色体の発達によりそれらはより体系化されてきた。
それらの源の集合体を意識と呼ぶが、その意識の進化種もまた様々である。
より三次元的に物事を捉える進化種、点、線、空間、多次、虚偽、幻影、幻想など実に様々なのだが、これらには全て一つの根源たる共通点をもとに派生する。
全体性の中で分離とも呼べる意識分野がごく少数存在するが、その中にもまた種類がある。
三次元的かつ精神的要素に傾倒した種、
完全に精神に傾倒した種。
これらの混じり合うこの世界において、その希とも呼べる種らは、通常種に紛れ、混合種とも似ても似付かぬ、まるで何らかの呪いをかけたらたかのようでもあり、圧倒的に偏ったとも呼べる精神に傾倒した種らは、基本的には現実的社会的問題ごとに対して、どれともにつかわない答えに至る。
それら以外の種にここ数年、特に混乱が濃厚さが増しているかのようにも見える。
現実社会を分断する二つの種や、スピ系界隈においても、情報は錯綜し、真偽に常に奔走しているかに見える。
これは間違いなく病的である。
その病の元凶…。
それこそはまさに情報とも呼べる。
遥か昔より、生命はある時、ある情報を元に、概念を想像し、定着させ、そこから意思を派生させ、進化に至る。
そして長い時間を費やし、情報的進化をする意識というものは肉体自体とは別に精神とそれぞれに意識を持たせた。
それをある意味強制的に統合させられた生命のうちの一種が勿論我々人類でもあるのだが。
この意識の基本形態は本来は肉体的、精神的と分断は存在しなかった。
やがて生命は偽りを生み出す事で分岐の歴史を迎え、時間や、様々たる要素の中でその進化種を増やした。
常識観念も然り、異端思想も然り、陰謀論も然りそれらは全てに個々的情報を元に真偽に脅かされ展開されている。
そしてそれは残念ながら実に遺伝子的(肉体的)といっても過言ではなく、
それを清算する為にこのsizeを書いているといっても過言ではない。
ただの情報としてではなく、
生命にもたらされた、
肉体派性型意識と精神派生型意識
の共生を真の意味でと思う。
だからこそこの意識の発達を現時点では洗練と表現する事は出来ないが、やがてはそうなることができればとの思いを込めている。
精神派生型意識とは永遠のズームアウトであり、全ての存在を、勿論悪と呼ばれるものも含め、一つの生態系的自然の流れの中でそれぞれの粒子の営み(衝突含め)として許されるべき、また愛されるべきものなのだと思う。
10億年前 多細胞の始まり
多細胞化第一要因 〜スプライジング
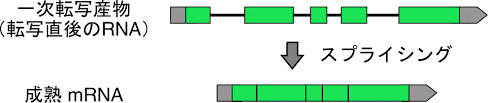
これまでの進化は放射線などの影響を主とする変異などの進化がベースで、これを茎進化という。
ここから行われるのは遺伝子の後輩や、種族の混合による冠進化がメインで動き出す。
これまではコドン配列をそのまま用いて染色体を形成していたが、いくつかの塩基配列をスキップすることで新しい染色体を作り出すと言う方法を身につける、勿論一つの遺伝子から効率的に多くのタンパク質をコードする為でもあるがそもそもの作り上げる染色体自体にもこの頃から変化が起きだす。
塩基配列の中には遺伝情報を持つ塩基配列(エキソン)とそれを持たない介在配列(イントロン)がある。
スプライジングは多細胞種から見られる反応で、多様な染色体情報を作り出す根源的な基盤となる。
しかしこのスプライジングされたコードはすぐに生命種に機能するわけでは無い、使用されるタンパク質が存在するしないでも変わり、このコード種は常にストックされていく、移りゆく変化の中での対応力として、事前に生命種が準備する。
なんらかの変異が外的環境に起きた時、まるで目覚めたかのように、その歯車は動いていく。
長い進化の歴史でイントロンは蓄積され塩基配列の長鎖を可能にした。
イントロンの存在は無駄なものではなく、言い換えるならば、その存在の意義は様々な場所に介在し、多様なエキソンパターンを構築する為だ。
だからこそ、それらを繋ぎ合わせる機能の入手が、爆発的な多様化の引き金となったのだ。
この長鎖の存在は、まるでそうなることをわかっていたかのようでもある。
多細胞化第二要因〜プラスミド
これまで生命は同じDNA情報からスタートし、様々な条件で枝分かれした
その反応は全てDNA配列パターンの変化による、
単細胞などは母体DNA以外のプラスミドと呼ばれる遊離DNAを持っている、
真核種には、酵母などに存在するが、それ以外にはほとんど含まれ無い。綺麗に分離できるようになったと言われればそれまでだが、当時の状況を考えると、生命がそれを利用しなかったとはとても考え難い。
ここで一旦整理したい、
基本として染色体情報は一貫して同じものを使い、
同じ情報を複製し続けてきたはずの生物の種に
これだけの多様性を与えてきたのは何なのか?
それがこの遊離プラスミドにある。
ねじ切る分裂の際、どうしても遊離するDNA情報らはその末端を隠せる機能が存在せず、膜内に存在する多様なタンパク質と結合していたと考えられる。
あるいは、新たにスプライジングされ出した、新種のタンパク質なども多く出現し、様々な反応が遊離プラスミドを通じ行われていた可能性はある。
真核種DNA末端にはメチル基が付属され安定しているが、それ以前には不安定さを残す、その不安定性はそのまま可能性とし利用するのが生命の持つ力でもある。
FtsZタンパク質と呼ばれる分裂タンパク質を原核種らは、使用する。
真核の多細胞種が扱うタンパク質は、より高次構造のものを扱う。
明確に作り込まれていないシステムという不安定性は、プラスミドと多様なタンパク質との反応をもたらした。
その結果単細胞種に新たな多様性をもたらし、多くの機能をもたらしたのだろう。
そして、この後に出てくる多細胞種の多機能性の由来、これはある意味細胞がそれぞれの分化の際、プラスミドを酵素的役割として利用した。
多細胞化第三要因〜減数分裂
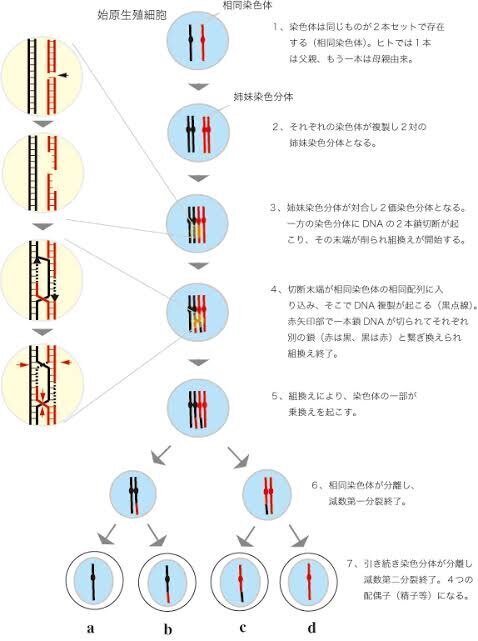
生殖細胞は、新たな、細胞周期サイクルを迎える。
この細胞は組み替えの後、体細胞同様の1度目の有糸分裂を起こす。
通常はここまでで一つの完成された染色体情報となる。
生殖細胞の場合2度目の分裂が起こる。
さらに別れ半分の染色体情報として組織化される鞭毛が生え、雌雄別々の生殖細胞を作り出す。
キアズマ:
染色体交差部分、タンパク質meikinが、働き出した事でこの交換作業は起こり出し、セントロメアという染色体接合部分の原動体からの分泌物、
ある意味セントロメアの進化の影響とも言える。
これらの母体たる生殖細胞には体細胞と違い半減期がなく、永続的に分裂を続け、その情報を延々と残していく。
その際作られる情報は、父、母2人分の染色体細胞から7兆通りを超えると言われており、ランダム性に富み、様々な、遺伝体型へと広がりを持つ。
そして、減数分裂を果たした種らは1/2の染色体を持ち寄り、一つの染色体の対を作り出す。
そうやって生み出されたのが、全能性を持つ受精卵や、胞子だった。
多細胞化第四要因〜幹細胞

この多様なプラスミドの中におそらく分化(分裂の際、自己複製+他の機能を持った細胞の複製)他機能細胞の能力を持つ機能の発生が隠されている。
単細胞から多細胞へと切り替わる際、プラスミドの総量の減少が見られる。
これはおそらく新たな機能を持つ多様な細胞形成の大元となった可能性がある。
幹細胞が生み出す異なる細胞の中に全能性細胞と多能性細胞がある。
全能性:全ての細胞になりうる細胞(受精卵のみ)
*全能性細胞も多能性細胞も作る事ができる。
多能性:胎盤などの生命を作る機関以外の細胞に分化する能力を持つ。
*全能性細胞にだけはなれない。
胚葉


生命の進化の中で
全能性を持つ胚細胞がある、始めからこれは、その状態で作られたわけではなく、細胞機能が複雑化される経緯の中で、徐々に作られた物。
最初は雌雄同体の種には漠然とした胚細胞が存在し、次に神経伝達のみを機能的に進めるニ胚葉目(外胚葉)が分化して作られる。
*ここで注意したいのは三胚葉全部を持っているのが全能性細胞だが、
生命の生成段階では勿論初めから全能性細胞などが存在したわけではない、単細胞スタートだから当たり前だ。
あくまでも原初の生体形成の流れにおいての流れは以下
単細胞→部分的減数分裂1回目→外胚葉生成成功→内胚葉生成成功→残された胚葉が中胚葉として進化する→減数分裂2回目導入→全能性の獲得
なのでこれは通常の減数分裂のサイクルと似てになるもの、数億年をかけて、様々な、コードとタンパク質の合成由来がこの時間の間にかかった。
なので、今の概念の多細胞化が突如起きたのでは無いということになる。
外胚葉:
多様なタンパク質は随時生み出されていく。
それと同時にスプライジングにより多様な塩基配列も常に生成される。
それらが始めて適応という相関関係を持った時、新たな細胞が作られ、組み込まれていく。
その流れで初めて生まれた他の機能が外胚葉、
ニュアンス的には皮膚や、神経、感覚など外部から侵入する電気系統的信号受信の生成でもある。
内胚葉:
やがて体細胞情報と遺伝情報を切り分ける三杯葉目が作られる。
要はこの時に単離した機能は、代謝に関与する細胞、消化、呼吸、尿路などである。
中胚葉:
後続的に単離したのが中胚葉(重要なのは、この時点で生殖細胞が含まれる事)
骨格、筋、液体循環、生殖器官(減数化)
後でも出てくるが、明確な減数分裂の発生もこの時ではないかと思う。
ついでに…。
実際には全能性細胞がどうやって生まれたかは知られていない、
小保方さんが作成したSTAP細胞は体細胞から幹細胞を培養する技術、
細胞に与える刺激によって細胞が変異する。
成功したと言ったが、その後色々あり、それは嘘だと認定されてしまった。
ES細胞とは人工で作られた、複製機能を持つ細胞。
受精卵から胚細胞を取り出し、分化後の細胞を作ることが出来る。
IPS細胞とは体細胞から多能性細胞まで還元に成功した細胞。
一見これは万能下に見えるが癌化のリスクがある。
STAP細胞の真相について細胞の進化について見ていきたい、そもそも細胞の活性化と進化の環境についてはあらゆる条件の合致の中でそもそも進化してきたものだ。
故にアナログな変化、
つまりは単純な環境変化においていくつもの分化を勧めてきたことは間違いない、
小保方さんが研究者としてではなく、あくまでも作業員としていくつもの刺激と変化の観察を行った事象の中に、
当時の環境変化との合致があったことは間違いない、
弱酸性の刺激と培養により、STAP細胞を発見したと言っている。
これはそのままその当時の海洋環境と近い、
可能性は確かに考えることが出来る。
そもそもであるが、このような新たな革新的結果というものは、これまでも権力によりいくつでも淘汰されてきた、
それ含め歴史のリフレインが起きていたと考えれることは自然の流れである。
実際、全能性細胞ともなれば、医療経済が黙ってはいない。
銅線を売るために葬られたフリーエネルギー、
純水が出回るのを阻止してまで添加水を出まらわせる理由、
陰謀論などではなく、
コレは既にはるか昔から受け継がれてきたものであり、
理研ですらアメリカのそういった力のもと成り立っている以上、例外ではない。
STAP細胞は、間違いなく存在していたと思われる、
明るみに成るかならないかもわからないが、
あまりにもコレは重複された歴史の悲劇を思い出させてしまう。
*生命種と染色体とゲノムサイズ(塩基数)
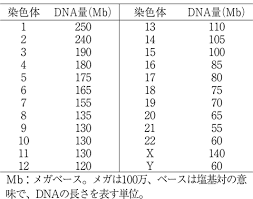

そもそもの話、現存する生命種は生き残ったかけら達、どのようなレベルの生命の段階であれ、それらは全て生き残った種。
過去にはおそらくは更に多くの種が存在し、淘汰され、発達段階にあった染色体らは、多くの阻害因子の元何度も書き換え、託されて今に至る。
単純に染色体の数で生命の進化段階がわかるはずもなく、ゲノムサイズですら全てではなくならざるを得ない。
染色体の量と塩基数と進化レベルからは単純に予測する事はできない。
そして、環境に適応できなければ、何らかの進化という意味で染色体は変動する。
消えたであろう、染色体の流れや、生命における謎は裸足でどこまで解消出来るのだろうか。
Y染色体の由来
一つの疑問が湧く、
何故Y染色体はあんなにも短いのか?
実際Y染色体の情報は、性別を決定するぐらいの情報しか記されていない、
また、脊椎動物のY染色体上部にのみSRY遺伝子というものが存在するが、これが存在するとするとその形状を保ち、男性へと機能していく。
とある説について
遺伝学者グレーブス博士の説
『生命種は進化の過程でY染色体が短くなっている、1400万年年後には地球上から全てのオスは消えるだろう。』
…これはそうはならない。
そう言い切れるのには幾つか理由がある、
哺乳類にY染色体を持たないにも関わらず雌雄が存在し、有性生殖を行う種がいる。
その種に関して言える事は、X染色体がSRY遺伝子を持っている。
それともう一つ大きな理由がある。
個人的にはこれが一番決定的な理由だ、
SRY遺伝子には癌抑制遺伝子が存在する。
正確にいうならSRYはそもそも癌抑制遺伝子として存在していた。
前回述べたように、染色体にはそれぞれ特有の癌に、対応する癌抑制遺伝子が存在する。
つまり、ある特定の癌抑制遺伝子を持った染色体が進化の中で不要な遺伝情報のみを削除し短くなり、必要な機能だけを残し、現在のY染色体の長さまで効率化させたと言える。
このようにして、
進化の過程では明らかに様々な理由のもと染色体は短くなっている、染色体情報も必要に応じ、不必要なら断裂したり、必要なら結合したり。
一昔前までは人とチンパンジーの遺伝子は、99.9%同じと言われていた。
しかし技術が進歩し、解析が進むごとに、実はその差は見えない程細部に入り混じり様々な違いが生じてくる。
それらは合成し、利用されるタンパク質に大きく影響を与える。
人類の染色体の数は変わりはないが、微細な変化はすでにスプライジングにより生まれてきているのだろう。
雌雄とは
Y染色体の経緯から考えると、ある意味男性は副産物なのだと思う。
そもそも性決定を引き起こす染色体は、xy以外にもzwがあり、虫、両生類、鳥などにそれらは使われており、やはり決定遺伝子がその中には含まれる。
副産物だと感じる所以はもう一つある、
減数分裂は2度分裂を起こすが、おそらくは、これはある時点から統合された機能で一本化された機能。
これまでの生命種の目的から考えると、やはり広義での優先事項と狭く深い目的がある。
減数分裂はあくまでも始まりは広義での多様化を優先とし、その中から染色体を効率化させ、かつ大きいゲノム数を保存できる種を作る、それが細胞自体の意思であり、全ては全体性のバランスを保つ為に形成されている。
勘違いしないで欲しいのは、人が選ばれしものだとは考えていない事だ、
個人的には責任を背負わされた種が、ある意味人なのだと思う。
スプライジングによりいくつもの情報が作り出され、多くの染色体へと派生し、多くの種を増やし、ある時期から雌雄同体が全てに与えられた、それでも続く度重なる組み替えの下、その中から減数分裂をある時期から組み込む種が現れる。
一度溢れかえった生命のエントロピーは、度重なる淘汰たる絶滅の末、現在の生態系を残すに至る。
*分類学の変化
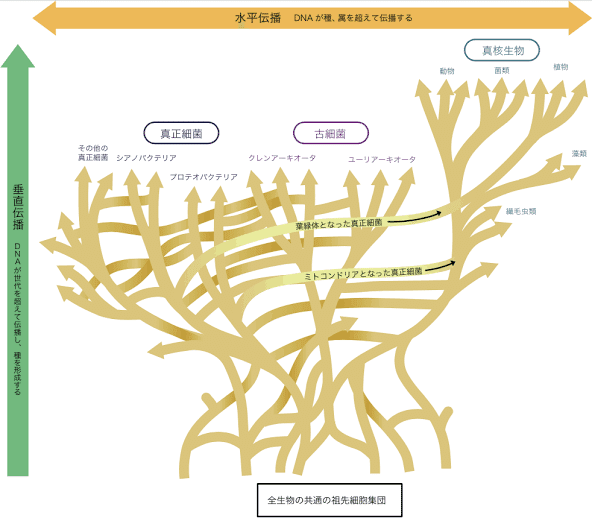
生物分類学ではリンネが分類階級をわけたがその後の多くの分野の活躍により、ミクロの視点から見れば、分子遺伝学がDNA塩基配列、細胞学が染色体、生化学がアミノ酸〜脂肪酸、発生学が胚葉や、幹細胞、解剖学が器官を、栄養学が全体の循環代謝を担い、分類学の発展に影響して来た。
また、生命種の発達経路には土壌の関連もある為、カンブリア機からは地質学、土壌学なども大きく関連してくる。
それらは全て電子伝達と、水の循環代謝の元全てが調和を保つからこそ、全現象学の網羅に至る。
分類学は、進化〜分岐〜表形(数量、割合)〜分子系統とその分岐方法も進化を続けてきた。
しかし現在ではその影響からこれまでの系統樹という考えでは当てはまらないケースが多く存在し出す、たとえば、イヌなどでさえ、系統で離れたり、離れていたはずの系統樹が実は近縁であったりと、実に様々な混乱が生じることとなった。
現在ではその影響から遺伝子の水平伝播が考えられ、多くの遺伝子の混在があったと考えられている。
これに大きく加担したのは、寄生虫、か、のみなどのつまりは遺伝子を運び出してしまうもの達の存在が影響する。
考えてみれば、それは確かにごく当然のことで、小さな要因が大きな変化をもたらす、まるでバタフライエフェクトや、カオス理論的な生命学と言えなくも無い。
記事を書く上で多細胞化要因を考えるのに長く時間を要した。
その理由はやはり系統の混乱にあった。
無性生殖から有性生殖が直接的に性別を意味するものではなく
全能性と多能性の細胞の経路はどこからか
幹細胞の分化が直接的には三胚葉に繋がらない
植物の祖は藻類であるにもかかわらず、多細胞系列にある時突如割り込んできたりと、
実に混乱をきたしているのが見てとれた。
結局何らかの小さな要因がこの混乱を解消する起点となると考えざるを得なくなり、
種に紛れた小さな遺伝子組み換え要因があるはずだと予測し、水平伝播、遺伝子持ち出しを掘り当てた感じとなった。
確実に少なくとも10億年も前に多細胞化の要因となる変化はもたらされ、カンブリア期(6億年前)まで生命の爆発が起きなかったとされてはいる。
勿論そのタイミングで起きた地上の変化は、酸素爆発と土壌の形成だが、
やはりその間引き起こされた2度の全球凍結と、現在では消滅したとされている、その間に起きたエディアカラ生物群。
データにすら残らない多くの生命種の発達と滅び去った多くの生態系が存在した可能性がある事は明らかだ。
マクロから見れば一つの流れのように見えるが、ミクロまでほじくりかえせば新たな事実が浮かび上がるのはどの世界を見ても事実である。
余談
生命の進化について、賛否両論ありますが…。
勿論一部では宇宙人が手を加えた説もあり、その中には信憑性のある物の存在も確かにある。
しかし、その情報で全てを片付ける前に、生命の神秘性、生命の持つ構造の中に存在する生命哲学を追求する事で、生命が存在する根本的な意味をむき出しにできるのでは無いかと思う。
別に地球外生命の関与の否定ではなく、一方関与の肯定でもなく、どこの段階で、どう様々が絡み合い今が存在するのか。
勿論超自然的構造、秩序の存在、神の力と言わんばかりの神秘性の存在、何らかの介入による何らかの変化…。
恐らくは生命の多様性以上に、全宇宙規模の多様性が本当のところは存在する、勿論文字として、言葉としてこれを知る術はない、
何らかの存在からのメッセージがあったとしてもそれが全てでは無い。
しかし一つだけわかっている事がある。
この世界は全てにおいてフラクタルである可能性が限りなくある事だ。
これがどう言う事なのか、勿論現存する情報全てが真偽に関わらず全て現存する事だ。
おそらくこれも全て含めてのフラクタルなのだと思う。
と言う事は真理のフラクタルが全てに撒き散らされている以上、それらを純粋に見極める事ができれば、全ての真実のみならず、真理を感覚的に捉える事が可能なのでは無いかと思う。
もしこれが人の脳で証明される物ならば、
もしそれが何らかの媒体で表現できる物ならば、
全人類が真理を感じ生きていく未来の可能性を
0から解き放つ事が出来るのでは無いかとも思う。
そう言う意味で私は一般的に人として生きる事はできなかったのかも知れない、でも多分、私が生まれた理由とその意味は、たったそれだけに尽きるのだと思う。
ヒドラ〜(まとめ)
多細胞、無脊椎、ニ胚葉種
ヒドラ
脳を持たずして眠る生命
果たして彼らの見る夢とはいったい何なのだろう
脳を定義する物
睡眠を定義する物
夢を定義する物
そんな我々に
そんな浅はかな我々に
何かを諭さんかのように
彼らは眠る
そうごく自然に
我々の考える定義など
まるで数多あるプラスミドの中の一つ
マクロたるプラスミドという概念は
かくも自然に
全てを包む母体の持つ可能性のように
秩序を突き動かし
生態系を生み出した
ヒドラは眠る
初めて神経伝達を切り離したその種らは
光と闇を選別し
意識を還す場を作り出す
夢とは確かに
現実に囚われる我々を
壊れる可能性のある精神を
そのバランスを保つための機能
しかし
ヒドラの見る夢はおそらく
我々のそれとは違う
深淵に還る為
眠りとは本来
真理と意識を繋ぐポータル
その乖離の狭間で
もがく我々に
優しく手を差し伸べるかの様な
ヒドラの眠り
まるでザラスシュトラのように
まるでアスクレピオスのように
感覚的にはヒドラに感じる感覚はこれ、
かく語りきのザラスシュトラは、これと同じくらい
美しく、か弱く、純粋性に富み、実際の彼もそうであったかに思う。
(ミクロとマクロを縦横無尽に泳ぐ為
おそらくその為に
あるじは僕にこれを見せたのだろう…。)
追記します。
若干意識が飛びすぎて何が言いたいのか具体性に欠けていたかに思う、それ程強いエネルギーに当てられてしまい、別に、具体性すらどうでも良くなるという症状に陥っていた。
単純に多細胞化は、多様性の中に一時的に埋もれ、明確な機能の産出には手を出さなかったという事で、遺伝情報の水平移動が頻繁におこり、当初有性生殖は局所的なものとして発言し、そのゲノム情報は、寄生や、微生物らが染色体情報を持ち出す事で伝播し。
その効率的機能から過半数の多細胞種の中で機能を働かせた。
これらは後に異化分裂と完全なるコピーを作る分裂細胞の共生へと繋がっていく。
この共生をもとに生み出されたのが幹細胞なるもの生命はまず神経系統を単離する事で眠るというサイクルを取り入れる、
これは言葉以上に実は面白い事で、明らかに実存の認識を推し量る機能の現れである。
その精査のためにこそ生命は睡眠を介しゆめをみるようになった。
やがて生命らは無性生殖から有性生殖へと明確なサイクルを生み出した。
それまで単一媒体の中で雌雄同体として生命を出現させていたそれらは更に広い意味での遺伝子混合案の一つとして0.5倍体の染色体を作り出し、種の淘汰を免れんが為の誓約にも近い
選択を選び取った。
これが俗に言う、減数分裂の起こりである。
生命種は強固なゲノム情報を作り出す為多くの繁殖方法を身につけた、
分裂から出芽に至り、栄養従属による繁殖、胞子、様々な選択がなされた、この流れの中には恐らく多くの問題が生じていた、群体らが癌細胞により、危機に晒されたのと同様、未完成な0.5媒体は多くの死滅にさらされながらも、その情報をRNAに蓄積させ、正当なタンパク質を身につけることに成功する。
これによって初めて生まれたものが全能性を持つ細胞だった。
全ての必要な情報を圧縮し、ゲノム内に押し込む。
解凍に必要な情報たるタンパク質を外部の環境から割り出すのに果たして何億年を有し、幾つの生命種の淘汰があったのだろう。
しかしそれらに一切の無駄はなく、全て全能性を持つ受精卵に生贄として捧げられた。
まさにそれは神の出現だったに違いない、
全ての生命は歓喜し、その神たる全能性細胞の元カンブリアの恵みの元生命はインフレーションを引き起こす。
SIZE〜10部〜HYDRA