コクヌストモドキの忌避行動 ~触角で感じる刺激~ 論文紹介

コクヌストモドキの忌避行動 ~触角で感じる刺激~
論文名 TRPA1‑mediated repellency behavior in the red flour beetle Tribolium castaneum
コクヌストモドキのTRPA1チャネルを介した忌避行動
著者名 Kenji Shimomura, Soshi Ino, Kazuya Tamura, Takehito Terajima & Motohiro Tomizawa
掲載誌 Scientific reports
掲載年 2022年
リンク doi.org/10.1038/s41598-022-19580-z
コクヌストモドキのTRPA1チャネルを介した忌避行動を調べた2022年の論文です。
コクヌストモドキは世界中に分布する米粉や小麦粉を食べる害虫であることから(漫画「コクヌストモドキ」参照)、害虫対策のために研究対象になっています。それだけではなく、ストレスに強いために飼育が容易で、卵から1ヶ月で成虫になるため、安定的に個体数を維持することができます。2008年には甲虫類では初めてゲノム解読が行われ、本論文でも使用しているRNA干渉によって標的遺伝子の発現をノックダウンすることができるため、分子生物学的研究を行うことも可能です。生態学的研究にも用いられており、甲虫類のモデル動物として、研究に使われています。
本論文で着目しているTRPチャネルは、熱や化学物質に反応して陽イオンを細胞内に流入させるチャネルです(漫画「TRPA1チャネル」参照)。TRPチャネルには多くの種類がありますが、有名なのはカプサイシンによって反応するTRPV1チャネルです。哺乳類では、このチャネルは43℃以上の熱や痛み刺激によっても反応して焼け付くような痛み(灼熱感)を引き起こします。つまり、ヒトではカプサイシンは熱や痛みと同じ刺激として認識されていることになります。また、高温だけではなく本論文で取り上げているTRPA1チャネルは、哺乳類では17℃以下の冷刺激に反応し、冷たい痛みを引き起こします。本論文では、他の昆虫の忌避行動に関連していたTRPA1チャネルに注目してコクヌストモドキの忌避行動を調べています。
「背景」にあるように、本研究は、SDGsの観点からコクヌストモドキに対する忌避剤として現在主流となっている合成忌避剤であるディートの代替品として植物に由来するエッセンシャルオイルを使用することを目的としています。本研究で取り上げているシトロネラールはシトロネラ油に含まれる成分です。主に石鹸用の香料や、他の香料を合成するための原料として利用されています。また、アロマオイルとしてそのまま使用されることもあります。天然成分ですので環境負荷が低いことが期待されます。コクヌストモドキは世界中に分布していることから、使用されている忌避剤の量は膨大なものになると考えられます。環境負荷の低い代替品が登場すればその影響は大きなものになるでしょう。本研究結果がその嚆矢になるかもしれません。
補足は論文には書かれていないことです。分かりやすくするために追加した文章になります。
この論文で分かったこと
TcTRPA1遺伝子をノックダウンしたコクヌストモドキでは、シトロネラールに対する忌避行動が有意に妨げられる。
TcTRPA1遺伝子は主に触覚で発現している。
触角を除去したコクヌストモドキでは、未処理体と比較して、シトロネラールの忌避活性が有意に減少する。
コクヌストモドキは触角に発現するTcTRPA1チャネルでシトロネラールを検出し忌避行動を起こすと考えられる。
[背景]
侵害受容とは、痛みや損傷の両方もしくはどちらか一方を引き起こす可能性のあり、逃避や忌避の行動パターンを引き起こす有害刺激を認知することです。侵害受容は、熱、機械的刺激、化学物質など、様々な種類の刺激によって引き起こされます。
刺激の検出には、複数の種類の膜たんぱく質、受容体、イオンチャネルが関与しています。一過性受容器電位(Transient receptor potential; TRP)チャネルは、脊椎動物と無脊椎動物で進化的に保存されている陽イオンチャネルです。TRPチャネルは、光、音、温度、味、化学・物理刺激など、様々な情報の主要なセンサーとして機能します。TRPファミリーは、構成する配列要素により、TRPC、TRPA、TRPV、TRPN、TRPM、TRPP、TRMLの7つのサブファミリーに分類されます。
TRPA チャネルは広く保存されており、N 末端領域には複数のアンキリンリピートが存在します。(補足:アンキリンリピートはアンキリンたんぱく質で見つかった33アミノ酸からなる繰り返し構造で、分子間または分子内の相互作用に関わる機能を持つ。)TRPA1チャネル は、侵害性熱刺激物質や有害な臭気物質・味覚物質などの刺激性化学物質によって活性化されます。哺乳類では、TRPA1チャネルは侵害受容神経に発現し、刺激性の天然化合物、アリルイソチオシアネート(AITC)、メントール、シンナムアルデヒドなど、多くの化学物質の侵害受容体として機能しています。(補足:アリルイソチオシアネートはマスタードやワサビの辛味成分、メントールは薄荷の香り成分、シンナムアルデヒドはシナモンの香り成分。)脊索動物のTRPAサブファミリーにはTRPA1しか存在しませんが、ショウジョウバエのゲノムには、脊索動物のTRPA1遺伝子の相同遺伝子であるTRPA1遺伝子、water witch遺伝子、pyrex遺伝子、painless遺伝子の4つのTRPAサブファミリー遺伝子が存在します。(補足:相同遺伝子とは配列の類似性から他の動物で見つかっている遺伝子と同じ遺伝子であると考えられる遺伝子のこと。)さらに、もう一つのTRPAサブファミリー遺伝子であるTRPA5遺伝子が甲虫類(鞘翅類)のモデル生物であるコクヌストモドキのゲノムに存在します。また、膜翅類のゲノムにはTRPA1遺伝子は存在しませんが、TRPA5遺伝子と膜翅類特異的TRPA(HsTRPA)遺伝子が含まれています。その他のサブファミリーについては、TRPMチャネルはメラニン色素細胞(melanocyte)から同定されたことから当初メラスタチンたんぱく質と名付けられました。さらに、哺乳類のTRPM8チャネルはメントールおよびその類縁化合物による化学刺激によって活性化されることが報告されています。TRPMサブファミリーはTRPM1から8までの8つのチャネルから構成されていますが、昆虫には1つのTRPM遺伝子のみが存在します。興味深いことに、ある種の天然由来のリガンドが複数のTRPチャネルを活性化することが示されています。例えば、メントールは哺乳類のTRPA1とTRPM8を活性化します。
植物由来のエッセンシャルオイルは、揮発性化合物の混合物で、二次代謝によって生成される環境保全型成分を含むことから、殺虫剤、忌避剤、産卵抑止剤として使用されています。(補足:二次代謝は生物の共通の生命現象に直接関与しない物質を合成する代謝のこと。)エッセンシャルオイルのうち、オガルカヤ属から抽出されるシトロネラ油は、数種の蚊、チャタテムシ、甲虫に対する強力な防虫剤として期待されています。シトロネラ油の化学成分のうち、モノテルペン化合物のシトロネラール(3,7-dimethyl-6-octenal)は、コクヌストモドキに対する殺虫活性および忌避行動を引き起こす主要化合物の一つです。
シトロネラール活性の分子認識機構については、これまでにショウジョウバエとハマダラカの2種の双翅類において、シトロネラールを忌避するためにTRPA1が必要であるという研究が行われました。また、アフリカツメガエルの卵母細胞に異種発現させた両者のTRPA1チャネルが、シトロネラールによって活性化されたことは注目すべき点です。
宿主植物に由来する匂いは、嗅覚神経の樹状突起上に発現する末梢性匂い受容体によって認識されます。昆虫の嗅覚受容体は、嗅覚受容体共受容体とともに陽イオンチャネルを形成し、嗅覚受容体共受容体は臭気感度の基準としてだけでなく、嗅覚受容体の局在と維持のためのシャペロンとしても機能します。(補足:シャペロンとは他のたんぱく質の機能を発揮するための立体構造の形成を助けるたんぱく質のこと。)そのため、嗅覚受容体共受容体遺伝子のノックダウンや無発現突然変異は、嗅覚機能の重大な喪失をもたらします。
ショウジョウバエでは、シトロネラール反応に2つの経路が関与しています。一つは嗅覚受容体による受容を介する経路で、嗅覚受容体共受容体遺伝子の変異により忌避活性が低下することが示されています。もう一つはGタンパク質共役型ホスホリパーゼCシグナル経路で、その下流でTRPA1チャネルが機能します。TRPA1チャネルのアイソフォームであるTRPA1(A)チャネルはシトロネラールによる味覚嫌悪を誘発します。(補足:アイソフォームは基本的機能が同じであるが配列の一部が異なるたんぱく質のこと。)また、ハマダラカやアカイエカなどの蚊の TRPA1 チャネルのアイソフォームである TRPA1sチャネル は、シトロネラールによって直接活性化されることが確認されています。他の昆虫種ではいくつかのTRPA1チャネルのアイソフォームが報告されていますが、コクヌストモドキについては、現在までのところ報告がありません。
TRPA1チャネルを介した化学物質受容に関連して、ミントから抽出したメントールがショウジョウバエの幼虫に侵害性回転行動を引き起こすこと、そしてTRPA1とTRPMが遺伝学的に相互作用し、嫌悪行動に必要なことが報告されています。培養細胞を用いた研究によって、ショウジョウバエのTRPA1チャネルはメントールによって活性化されないことがわかり、TRPA1チャネルがメントールの認識に機能するかどうかについての議論が続いています。
しかし、シトロネラールの甲虫に対する忌避メカニズムはまだ解明されていません。また、コクヌストモドキにおいて l-メントールがTRPMチャネルを介した接触忌避行動を誘発することを以前に報告しましたが、メントールが TRPA1チャネルを介した忌避行動を誘発するかどうかは分かっていません。本研究では、RNA干渉によるTRPA1遺伝子のノックダウンにより コクヌストモドキのシトロネラールに対する忌避行動を調べ、TRPA1が忌避反応に関与しているかどうかを明らかにすることを目的としました。(補足:遺伝子ノックダウンは遺伝子の転写物であるmRNAからたんぱく質に翻訳されないようにすることで、たんぱく質の機能を阻害すること。図1参照。)さらに、シトロネラールに対する嗅覚受容体を介した忌避行動を調べ、TRPMチャネルとの組み合わせでメントール由来の忌避行動にTRPA1チャネルが関与することを明らかにしました。

[結果]
シトロネラールに対する忌避行動
区域選好試験は、半分を忌避剤で処理したろ紙を敷いたガラス皿に、20匹のコクヌストモドキ入れ、3時間後にろ紙の忌避剤処理区域と対照区域それぞれにいるコクヌストモドキの数を計測しました。コクヌストモドキの数から忌避値を算出し、5回の実験の平均値を求めました。忌避値は-1から1までの値で、全てのコクヌストモドキが対照区域にいる場合(忌避した場合)に1、忌避剤区域にいる場合(選好した場合)は-1となり、各区域で同数の場合は0となります。
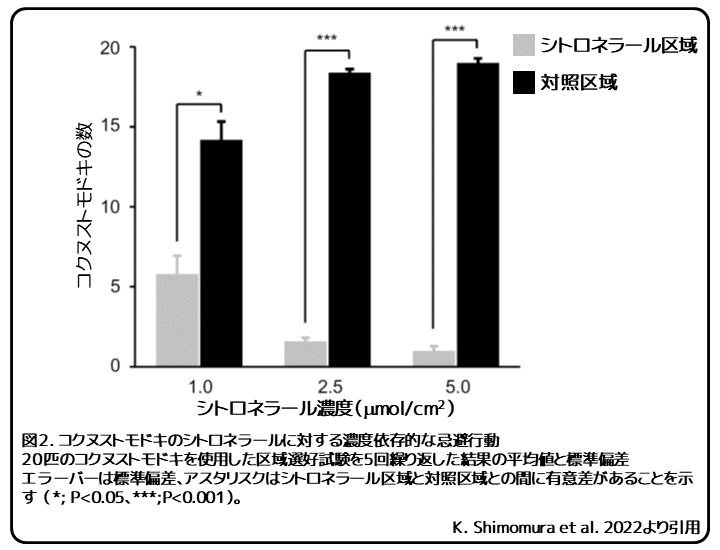
この区域選好試験によって、コクヌストモドキのシトロネラールに対する濃度依存的な忌避活性が明らかになりました(図2)。シトロネラールは2.5μmol/cm2以上で高い忌避活性を示しました。この結果に基づいて、以降の実験のシトロネラールの濃度を2.5μmol/cm2に設定しました。
シトロネラールの標的を明らかにするために、コクヌストモドキのTRPA1(TcTRPA1)遺伝子をクローニングし、全長アミノ酸配列が、他の甲虫類のTRPA1チャネルと高い配列相同性を持つことを明らかにしました。系統樹解析によって、昆虫のTRPA1チャネルの中で甲虫のTRPA1チャネルはクラスターになっていることが分かりました。
RNA干渉による標的遺伝子のノックダウンは、量的逆転写PCR(quantitative reverse-transcription PCR; qRT-PCR)によって評価しました。TcTRPA1遺伝子に対する二重鎖RNAを蛹に注入することでTRPA1遺伝子の転写を抑えることに成功しました。標的であるTcTRPA1遺伝子の転写物は27%にまで減少しました(図3a)。対照群である水とEGFP遺伝子に対する二重鎖RNAの注入では、注入していないコクヌストモドキと比較して有意な差は見られませんでした。
TcTRPA1遺伝子をノックダウンしたコクヌストモドキの行動実験から、忌避行動の有意な変化が明らかになりました。TcTRPA1遺伝子のノックダウンによって、シトロネラールに対する忌避行動が有意に妨げられました(図3b)。水やEGFP遺伝子に対する二重鎖RNAを注入したコクヌストモドキへの忌避活性は、注入していないコクヌストモドキと比較して有意な差はありませんでした。
TcTRPA1遺伝子をノックダウンしたコクヌストモドキを、標準的な合成忌避剤であるディート(N,N-diethyl-3-methylbenzamide)を使用した区域選好試験で比較しました。TcTRPA1遺伝子をノックダウンしたコクヌストモドキはディートに対して忌避行動を示しましたが、注入していないコクヌストモドキとの有意な差はありませんでした。

TcTRPA1遺伝子転写物の相対的発現レベルと忌避行動での触覚の関与
いくつかの組織でのTcTRPA1遺伝子転写物の相対的発現レベルをqRT-PCRによって解析しました。TcTRPA1遺伝子は主に触覚で発現しており、その発現レベルは腹部での発現の34倍以上でした(図4a)。
触角に発現するTcTRPA1チャネルによってシトロネラールが検出されていることを明らかにするために、触角を切り取ったコクヌストモドキを使って区域選好試験を行いました。触角を除去したコクヌストモドキは未処理体と比較して、忌避値が有意に減少しました(図4b)。

RNA干渉によるコクヌストモドキの嗅覚受容体共受容体(TcOrco)遺伝子のノックダウン
触角によるシトロネラール検出の特異性を解析するために、TcOrco遺伝子に対する二重鎖RNAの注入を行いました。qRT-PCRの結果に基づくと、注入されたコクヌストモドキではTcOrco遺伝子転写物が25%にまで減少しました(図5a)。続けて、区域選好試験を行ったところ、シトロネラールの忌避活性は注入していないコクヌストモドキと有意な差は見られませんでした(図5b)。
いくつかの組織でのTcOrco遺伝子転写物の相対的発現レベル解析から、TcOrco遺伝子は主に触角に発現していました。

l-メントール忌避でのTcTRPA1チャネルの関与
RNA干渉を用いた実験により、l-メントール忌避でのTcTRPA1チャネルの関与を明らかにしました。TcTRPA1遺伝子をノックダウンしたコクヌストモドキでのl-メントールの忌避活性は、注入していないコクヌストモドキと有意な差は見られませんでした。しかし、TcTRPM遺伝子をノックダウンしたコクヌストモドキと、TcTRPM遺伝子とTcTRPA1遺伝子を同時にノックダウンしたコクヌストモドキは、注入していないコクヌストモドキと比べて忌避値が有意に減少することが分かりました(図6)。

[考察]
昆虫では、TRPA1チャネルは広く発現しており、有害な熱や刺激性化学物質によって活性化されます。コクヌストモドキのシトロネラール嫌悪の分子メカニズムを明らかにするために、RNA干渉によるTcTRPA1遺伝子のノックダウンを行い、忌避行動を評価しました。本研究結果から、区域選好試験でTcTRPA1遺伝子をノックダウンしたコクヌストモドキはシトロネラールによって忌避されないことから、シトロネラールに対するTcTRPA1チャネルを介した忌避行動が明らかになりました。
合成忌避剤であるディートは、絶対的な忌避剤であり続けています。ディートによる忌避には複数の経路とメカニズムが関与していますが、明らかにはなっていません。以前の研究で、コクヌストモドキはディートに対する忌避行動を示しました。しかし、知る限りでは、ディートが昆虫のTRPチャネルを活性化することを報告した研究はありません。本研究では、TcTRPA1遺伝子ノックダウンを行いディートに対する忌避行動を区域選好試験で解析したところ、TcTRPA1遺伝子をノックダウンしたコクヌストモドキの忌避値は、未処理のコクヌストモドキと有意差がなかったことから、TRPA1チャネルとシトロネラールは特異的に相互作用している可能性が示されました。(補足:ディートはTRPA1チャネルを介しておらず、TRPA1チャネルを介した忌避行動はシトロネラール特異的である可能性が示された。)
成虫のいろいろな部位でのTcTRPA1遺伝子の発現解析から、主な発現部位が触角であることが分かりました。同様の結果が半翅類であるトビイロウンカで示されていました。これらの結果から、本研究では、コクヌストモドキは触角による受容によってシトロネラールを忌避すると考えました。触角を切り取り、未処理のコクヌストモドキと忌避値を比較したところ、有意な減少が示されました。一方で、これまでの研究ではl-メントールの接触忌避活性は、触角ありとなしの間で有意差はありませんでした。これらの結果から、シトロネラールは触角に発現するTcTRPA1チャネルを介して忌避行動を引き起こすことが明らかになりました。
双翅類では、Gたんぱく質/ホスホリパーゼCシグナル経路でのTRPA1チャネルの間接的な寄与と、直接的な嗅覚受容体共受容体の寄与の2つがシトロネラール忌避のための経路として、ショウジョウバエで明らかになりました。ミカンコバエでは、Orco遺伝子のノックダウンによって、シトロネラールによる産卵忌避の減少が起きました。ハマダラカでは、シトロネラールは直接TRPA1チャネルに結合します。鱗翅類では、アフリカツメガエルの卵母細胞に発現させたオオタバコガのTRPA1チャネルが熱センサーだけでなく、シトロネラールを含む刺激性化学物質センサーとしても機能することが分かっています。
対照的に、半翅類の場合は、アカヒゲホソミドリカスミカメで、TRPA1遺伝子ノックダウンはシトロネラール誘導性の逃避行動を減少させましたが、Orco遺伝子ノックダウンは反応に変化を起こさなかったことから、シトロネラール忌避のための嗅覚受容体非依存的経路が考えられました。甲虫類の嗅覚受容体を介したシトロネラール忌避を明らかにするために、本研究ではTcOrcoを介したシトロネラール忌避を確かめました。区域選好試験によって、TcOrcd遺伝子をノックダウンしたコクヌストモドキの忌避行動は、ノックダウンしていない場合と比べて有意差がないことが明らかになりました。嗅覚受容体共受容体は広く共受容体と共発現し、昆虫全体に強く保存されているため、これらの結果から、コクヌストモドキの嗅覚受容体によるシトロネラールの受容システムの非介在性が考えられました。ここまでの結果を総合すると、昆虫類でのシトロネラールの受容には進化的多様性があると考えられます。
複数のTRPチャネルがメントールの受容に関わっていると考えられています。ショウジョウバエの幼虫では、メントールはTRPMチャネルとTRPA1チャネル依存的に多樹状クラスiv侵害受容神経を必要とする侵害性回転行動を引き起こします。その解析では、TRPM遺伝子とTRPA1遺伝子のそれぞれのホモ変異体の幼生がメントールに対して有意に少ない回転行動を示しました。さらに、TRPM遺伝子とTRPA1遺伝子の両方のヘテロ変異体の幼生では、メントールに対する回転行動は有意に抑制されたことから、メントール感受での遺伝的相互作用が分かりました。対照的に、コクヌストモドキ成虫のメントールに対する忌避行動はTcTRPMチャネルによって影響を受けました。TcTRPM遺伝子の相対発現レベルは、腹部、胸部、頭部、脚、触角で僅かな差しかありませんでした。しかし、本研究では、TcTRPA1遺伝子は主に触角で発現していましたが、触角を取り除いた成虫はl-メントールに対する忌避行動をまだ示したことから、メントールによって引き起こされる忌避にはTcTRPMチャネルだけが必要であると考えられます。ほ乳類の場合では、TRPチャネル以外に、メントールは異なる種類の電位依存性イオンチャネルとγ-アミノ酪酸受容体、グリシン受容体、ニコチン性アセチルコリン受容体のようなリガンド依存性イオンチャネルを作動させることから、コクヌストモドキ成虫のメントールによる忌避行動にはTRPチャネルとは異なる受容体が存在すると考えられます。
本研究結果から、甲虫類のTRPA1チャネルが持続可能な昆虫忌避剤の標的となる可能性が考えられます。興味深いことに、ショウジョウバエのTRPCサブファミリーに属するTRPγが、忌避行動を開始するためにシトロネラールによって直接活性化されることが最近報告されており、甲虫類でもさらなる実験とスクリーニングが望まれます。
いいなと思ったら応援しよう!
