
微生物による窒素固定の謎めいた進化:地球の過去からの洞察

メインコンテンツへスキップ
微生物学の動向
ログイン
検索...
レビュー|オンライン公開中
微生物による窒素固定の謎めいた進化:地球の過去からの洞察
https://www.cell.com/trends/microbiology/fulltext/S0966-842X(23)00091-4

ホリー・R・ラッカー
ベテュル・カサール
オープンアクセス掲載:2023年4月13日DOI:https://doi.org/10.1016/j.tim.2023.03.011
PlumXメトリクス
ハイライト
生物学的窒素固定の鍵となる酵素であるニトロゲナーゼは、進化的に特異な存在である。というのも、大気中の窒素を固定窒素に還元する触媒機構は、進化的に1つしか知られていないからである。
ニトロゲナーゼは3つの金属アイソフォーム(鉄、モリブデン、バナジウム)を持ち、そのため金属の利用可能性は生物地球化学的サイクルに依存している。
モリブデンは現代の海洋では豊富に存在するが、太古の海洋では非常に希少であり、現代の陸上環境では依然として希少である。その希少性にもかかわらず、初期のニトロゲナーゼは、豊富な鉄よりも触媒作用に優れているモリブデンを最初に利用したと提唱されている。
窒素循環の進化と多様化を理解するためには、まず生物学的窒素固定を生み出す古生態学的・惑星学的条件を理解する必要がある。
要旨
バイオマスの合成に窒素が不可欠であることを考えると、窒素固定の進化が生物圏のほぼ全域に変化をもたらしたことは間違いない。今日に至るまで、どのような惑星的条件が窒素固定を生み出したのか、またこれらの条件がどのように進化的に窒素固定を維持してきたのかについて、統一された見解はない。興味深いことに、ニトロゲナーゼが機能するために必要な金属の濃度は、地球の歴史を通じて変化してきた。本総説では、金属と窒素のサイクルとニトロゲナーゼの進化との相互関連、および現代の窒素サイクルの形成における古代の生態学の重要性について述べる。窒素サイクルの深い過去を探ることで、窒素の利用可能性を中心とした人類の当面の環境問題に対する洞察が得られると主張する。
キーワード
窒素固定
ニトロゲナーゼ
金属アベイラビリティ
分子古生物学
惑星微生物学
本文
回転する時計の歯車のように、地球上のさまざまな生物地球化学サイクルは互いに連動している。その結果、ある生物地球化学的プロセスの継続が、別の、しかしつながっているサイクルの進行によって影響を受けることがある。このようなフィードバックは、窒素循環で観察することができる。窒素は核酸やタンパク質の基本単位であり、生命にとって不可欠である。しかし、窒素の最大の貯蔵庫である大気中の窒素は、高エネルギー現象によって解離する少量の窒素を除いては、ほとんどの生命にとって窒素源として利用することができない。ニトロゲナーゼは、窒素をより生物学的に利用しやすい形態であるアンモニアに固定することができる唯一の酵素であり、陸上と海洋の両方の環境における窒素循環の中心的な役割を担っている。そのため、あらゆる環境に存在するすべての生物は、窒素固定によって生成される固定窒素を、直接的または間接的にニトロゲナーゼに依存している。
ニトロゲナーゼを介した生物学的窒素固定(BNF)は、地質学的な時間スケールで栄養素としての窒素の性質が限られているため、拡大する地球生物圏において長い間重要な役割を担ってきた [2.]。窒素肥料生産における固定窒素の制限を工業的に回避するハーバー・ボッシュ・プロセスは、30億年前の分子革新を補うためのエネルギー的にコストのかかる試みである [4.]。金属酵素であるニトロゲナーゼは、特定のアイソザイムによって、モリブデン、バナジウム、または鉄のみを取り込む、いくつかの鉄-硫黄クラスターを利用する [5.]。BNFの進化を決定づけた地球の歴史の要因は、まだほとんど分かっていない。例えば、最初のニトロゲナーゼがどのように進化したのか、初期の窒素固定が現存するプロセスとどの程度異なっていたのか、代替金属を利用するためにどのような選択圧が必要だったのか、現存する生物の窒素固定の配列の多様性は過去の生態学的条件の変化をどの程度示しているのか、まだ分かっていない。
現在のところ、どのような惑星的条件が窒素固定を生み出したのか、あるいはこれらの条件が進化的にどのように窒素固定を維持してきたのかについて、統一的な見解はない。しかし興味深いことに、窒素固定酵素が依存する金属の濃度は、地球の歴史を通じて変化しており、環境によっても異なる [6.]。本総説では、(i)関連する金属の時間的・空間的利用可能性、(ii)微生物窒素固定とその様々な金属アイソフォームの起源、(iii)現代のマルチバイオーム占有窒素循環の形成における古代の生態学の重要性、について論じることで、金属と窒素の循環とニトロゲナーゼの相互関係、およびニトロゲナーゼを介した窒素固定が惑星に与える影響について述べる。最後に、窒素サイクルの深い過去を探ることで、窒素の利用可能性を中心とした我々の身近な問題に対する洞察が得られるかもしれないと主張する [7.,8.]。
初期の地球と潜在的金属補因子の利用可能性
N2ガスが現在の地球大気を支配しているように、N2も先カンブリア時代の大気中に非常に豊富に存在していたが、おそらく現代の値よりも低い分圧であった [9.,10.]。ニトロゲナーゼを介したBNFの進化以前は、窒素の固定型は、N2の三重結合を切断するのに高いエネルギーを必要とする生物反応によってのみ生成できた(式1)。生物反応は、N2を還元するために、雷、流星衝突、またはFe-S鉱物触媒を用いた紫外線光触媒によるエネルギーを使用していたであろう [11.,12.,13.]。これらの反応だけでは、生命が進化し拡大し始めるにつれて増大する固定窒素の需要を満たすことができなかった可能性が高いため、N2を還元するための生物学的メカニズムの進化に対する選択的圧力となった [13.]。
�
2
+
8
�
+
+
8
�
→
2
�
�
3
+
�
2
[1]
窒素酸化酵素が惑星規模の生命に与える影響を理解するには、その出現と進化を形成したと思われる大気条件やそれによる金属の利用可能性など、初期の地球の条件を理解する必要がある [14.]。古生代の堆積物から得られた窒素同位体比を用いて、ニトロゲナーゼは32億年前に誕生したと推定されている [15.]。この推定では、ニトロゲナーゼ、あるいはニトロゲナーゼに似た祖先の起源は、大酸化現象(GOE)以前の古生代に位置づけられる。GOE以前、大気にはほとんど酸素がなかったが、GOEの数百万年前には、浅い海洋環境に少量の酸素が蓄積し始めていたはずである [16., 17., 18.]。
大気中の酸素が増加する前、古代の外洋の状態は著しく還元的であり、そのためFe2+の形をした鉄が、10-7 mMの範囲の濃度で、溶存成分として豊富に存在していた [19.]。古生代のほとんどの期間、Fe2+の濃度は水柱全体で高いままであった。酸素性光合成が進化し始めると、酸素が生成され、その結果、主に表層付近でFe2+がFe3+に酸化される「渦」が発生した [16.]。GOEは、その名の通り、海洋の酸化還元化学をより酸化された状態にシフトさせた。Fe3+は水に非常に溶けにくい鉄(III)酸化物を形成するからである。したがって、ニトロゲナーゼの起源がGOE以前にさかのぼるということは、生物学的に利用可能な鉄が豊富な環境で酵素が進化したことを意味する。このように、鉄はニトロゲナーゼ複合体の不可欠な部分であり、鉄-硫黄クラスターの形と金属補酵素の形の両方の成分に含まれている。ニトロゲナーゼのアイソフォームのひとつである鉄ニトロゲナーゼでは、金属補酵素は鉄のみを含む。
鉄とは異なり、モリブデンはアルケアン海には特に少なかった。アルケアンにおけるモリブデンの供給源のひとつは、海底熱水噴出孔であった可能性がある [20.]。酸素レベルが上昇するにつれて、地上の硫化鉱物の酸化的風化によって、これらの鉱物に濃縮されたモリブデンが河川に、そして最終的には海洋に動員されたであろう [21.](図1)。モリブデンは現在、海洋で最も豊富な微量金属である [22.]。GOE以前はモリブデンの濃度が著しく低かったにもかかわらず、推定される祖先の塩基配列による系統学的再構築によって、最も初期のニトロゲナーゼは、V-またはFe-ニトロゲナーゼではなく、Mo-ニトロゲナーゼ(Nif)であった可能性が高いことが明らかになった [23]。バナジウムは、現代の地球上ではモリブデンよりも存在量が少ないが、古生代の海洋では、より酸性度の高い環境で生物学的に利用可能であった [24.]。今日、バナジウムは海洋で2番目に豊富な微量元素である [25.]。
図1
図1地球地球化学と窒素固定の年表。
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
ニトロゲナーゼの起源
固定された、あるいは生物学的に利用可能な窒素は、生命にとって極めて重要であり、ヌクレオチドやアミノ酸などの重要な分子に含まれている。そのため、窒素のアンモニアへの変換を触媒する酵素は、生命の進化の歴史において重要な技術革新であった。ニトロゲナーゼは、生命が重要な代謝反応を触媒できる唯一の酵素を進化させたという、進化的特異点の一例である [26.]。ニトロゲナーゼが自然淘汰の圧力によって進化したのか、あるいはニトロゲナーゼの起源が確率的偶然や歴史的偶発的偶然によるものなのかは、いまだに不明である。
ニトロゲナーゼの起源に関する最近の古分子解析によると、ニトロゲナーゼは、N2を固定できないマチュラーゼのような前身(アセンブリー足場とも呼ばれる)から生まれたことが示唆されている [23.,27.]。マチュラーゼのNifENタンパク質は、成熟した金属鉄-硫黄クラスターを形成するための足場として働き、それがNifD活性部位でニトロゲナーゼ複合体に移行する。マチュラーゼ前駆体がモリブデンを取り込むことができたかどうか、もしできたとすれば、そのようなプロセスが現存するNifENで観察されたのと同様のメカニズムで起こったかどうかは不明である。また、最初のニトロゲナーゼやマチュラーゼ前駆体が、生命のどの領域で誕生したのかについても議論がある。以前は、ニトロゲナーゼは古細菌に起源があるとされていたが [28.]、最近の研究では、最古のニトロゲナーゼは細菌の祖先に存在した可能性があると推測されている [29.]。
謎に包まれたニトロゲナーゼの起源を解読し、金属選択に関する疑問を探るために使われ始めている手法のひとつが、祖先配列再構築(ASR)であり、タンパク質の塩基配列を過去に遡って追跡するために利用されている [27.,30.]。ASRは進化仮説を研究するための貴重なツールであるが、生存者バイアスによる特定の配列の過大評価を考慮することが重要である [31.]。古代タンパク質の再構築は、基本的に生存子孫酵素の祖先を再構築することに限定される。例えば、ニトロゲナーゼの基質チャンネル残基の最近のASRは、推定されるニトロゲナーゼの祖先は、現存する配列とは異なる基質チャンネル配列を持っていた可能性が高いことを示している[32.]。これらの残基の違いは、初期のニトロゲナーゼがHCNのような代替基質を還元するための選択圧力の結果かもしれない。したがって、現存する配列と現在の酵素活性だけに基づく配列解析の進化的解釈には慎重であることが重要である。ASRアプローチの威力は、現存する酵素が占める配列空間を超えて機能的研究を拡張し、進化の歴史を通して機能的変異のパターンを特定することにある [30.,33.]。
ニトロゲナーゼが進化したとき、おそらくマチュラーゼ様タンパク質から進化したのであろうが、その進化はおそらくモリブデンを利用した後に、代替のFe-またはV-コファクターを組み込んだのであろう。系統学的分析によると、最古のニトロゲナーゼの祖先はモリブデン依存性と一致する配列特徴を示している [23.,34.]。地質学的証拠(同位体分別値など)も、GOEよりかなり前のアルキア紀中期までさかのぼるモニトロゲナーゼの初期の存在を支持している [15.]。代替的なニトロゲナーゼは、古代の堆積物には見られない、はっきりとした非常に負のδ15Nシグネチャーを持っている [15.]。さらに、全てのジアゾ栄養細菌はモニトロゲナーゼを持っているが、代替ニトロゲナーゼも持っているのはごく一部である。
なぜモリブデン、バナジウム、鉄がニトロゲナーゼの金属補酵素として選択されたのかについては不明な点が多い。GOE以前はモリブデンが希少であったにもかかわらず、モニトロゲナーゼの動力学的優位性が、より豊富だが効率の悪い代替金属よりも、補酵素としてのモリブデンの使用を支持したのかもしれない。より一般的であることに加え、Mo-ニトロゲナーゼはより効率的(すなわち、N2 1 molあたりに必要なATPがより少ない)であり、また、より速い速度で還元反応を触媒する [35.,36.]。現代の窒素固定をまだ支えている最小モリブデン濃度閾値に関する知識は、鉄よりもモリブデンがどのように最初に補酵素として選択されたかを明らかにするかもしれない。フィリッピ(Philippi)らは、無酸素状態の原生代の類似系であるカダーニョ湖における低モリブデン濃度の影響を調査し、紫硫黄細菌が、モリブデンの利用可能量が限られている(<10 nM)にもかかわらず、モニトロゲナーゼを独占的に使用していることを発見した [37.]。バナジウムが原核生物のメタロームで最も利用されていない金属の一つであることを考えると、バナジウムが補酵素として利用されていることは驚きである [30.]。バナジウムは今日、豊富な微量金属であるが、深い時間のバナジウム濃度を特定することは困難であり、その結果、バナジウムがアルケアン時代に生物学的に利用可能であったかは不確かである [24.]。未解決の疑問は、なぜニトロゲナーゼが、マンガン、コバルト、ニッケルよりもバナジウムを利用したのかということである。
なぜ特定の金属が他の金属よりも選択されたのか、また他の金属因子がどの程度使われた可能性があるのかをより良く理解するためには、実験室内で古代の地球条件を再現する [38.]だけでなく、古代の地球の環境アナログを使う [39.]ことによって、元素の選択を決定する進化のメカニズムを解読する必要がある。これらの選択法則は、タンパク質や機構レベル、さらには生物レベル、さらには生物地球化学サイクルレベルにまで影響を及ぼす。したがって、窒素固定など、これらの元素を利用する代謝の進化を考える際には、元素の選択と元素量の経時的変化を組み込むことが重要である。例えば、アルケアン海洋のバルク金属濃度が、熱水噴出孔[20.]のような、最古のニトロゲナーゼが出現したと思われる局所的ニッチの金属存在量を正確に反映しているかどうかは不明であり、したがって考慮することが重要である。過去において進化がどのようにニトロゲナーゼを形成してきたかをより良く理解することで、この酵素が将来どのように進化し続けるかを理解するためのより良い準備が整う。
現代の窒素循環
過去からの洞察
ニトロゲナーゼによる窒素固定が始まった時代と、それが生じた環境条件が大きく異なることを考えると、現代の窒素固定が古代のニトロゲナーゼの進化によってどのように形作られてきたかを考えることは興味深い。太古の過去が現代のプロセスをどのように規定してきたかをよりよく理解するためには、いくつかの未解決の疑問がある。第一に、BNFが無酸素状態で進化したことを考えると、何がこのプロセスを現代の様々な酸化的環境で増殖させることを可能にしたのか?第二に、代替的なニトロゲナーゼの進化によって、ジアゾ栄養細菌は、これらのアイソザイムがなければアクセスできないようなニッチへの進出をどのように可能にしたのだろうか?最後に、シアノバクテリアのような現代の窒素固定の大部分を担っている微生物は、いつ、どのようにして初めてニトロゲナーゼを手に入れたのだろうか?これらの疑問を念頭に置きながら、現代の窒素固定について、特に生息環境、金属利用、酸素や栄養塩の制限などの環境ストレス要因への適応に焦点を当てて議論する。
海洋環境
窒素固定は、現代の海洋において依然として重要なプロセスである。陸上環境と比較して、海洋環境における窒素固定はいくつかの理由からよく研究されている。第一に、海洋は地球表面の70%以上を覆っており、窒素固定と窒素循環全体の影響を研究する上で、はるかに大きなバイオームとなっている。また、海洋では、数センチメートルで物理化学的環境が劇的に変化する土壌生物群集に比べ、混合によってより大規模な均質性が形成される。N2は海洋における窒素の支配的な形態であり(94%)、その他の窒素種は相対的に非常に少ない(6%) [40.]。これらの二次的窒素種のうち、最大の貯留量は硝酸塩の形の固定窒素(88%)であり、溶存有機窒素がそれに続く。粒子状有機窒素、亜硝酸塩、アンモニアはすべて、海洋環境では非常にマイナーな窒素種である(1%未満)。一般に、海洋環境も陸上環境も窒素源が不足しており、これらの環境では生物学的に利用可能な窒素源を作り出すジアゾ栄養細菌の役割がより重要になる [41.]。
シアノバクテリアのトリコデスミウム属は、おそらく海洋生態系で最も重要な窒素固定生物である。海洋において窒素固定によって年間100-200Tgの反応性窒素が生産されるが、トリコデスミウムは年間60-80Tgの窒素を供給していると推定されている [42.]。現代の海洋が酸素化されていることを考えると、トリコデスミウムのような好気性窒素固定生物は、酸素からニトロゲナーゼを保護するメカニズムを採用しなければならない。他の多くのシアノバクテリア属とは異なり、トリコデスミウム属はヘテロシストを持たず、むしろジアゾサイトと呼ばれる細胞のゾーンを介して酸素からニトロゲナーゼ酵素を保護する [42.]。このジアゾサイトには、ヘテロシストを取り囲む糖脂質層がないため、細胞内へのガス流入を増加させることができる [43.]。トリコデスミウムは、O2フラックスの増加の代償として、窒素固定のためにそのジアゾサイトへのN2フラックスの増加とトレードオフを行うことができる。しかしながら、最近のモデルでは、トリコデスミウムがジアゾサイトの空間的分離を必要とせずに、特に蓄積された固定炭素の呼吸による保護によって、細胞内のO2レベルを低く維持できることが示された [43.]。細胞内の酸素からニトロゲナーゼを保護するこのメカニズムに加えて、トリコデスミウムの窒素固定は時間的に制御されており、酸素生産が低下する日中に最大の活性が生じる [42.,44.]。
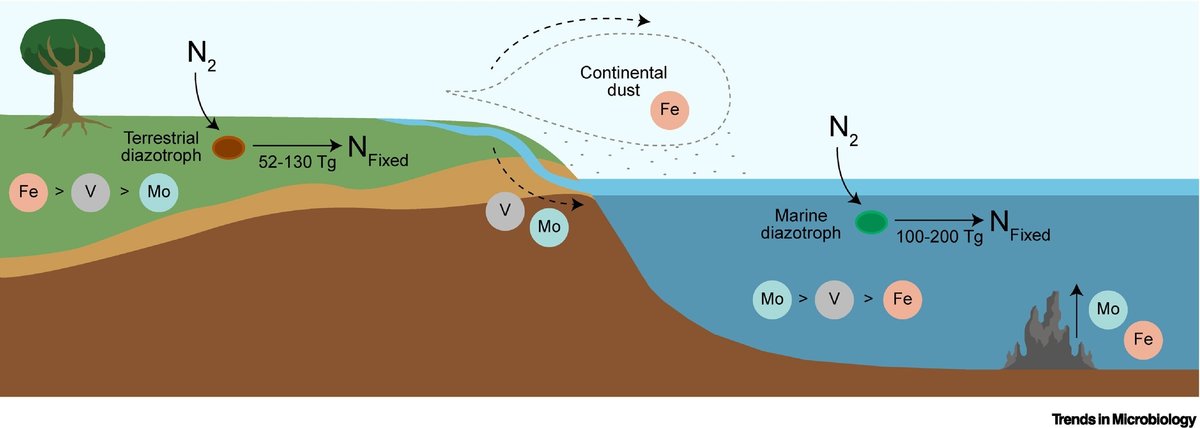
酸素阻害に対処する戦略を開発しなければならないことに加え、トリコデスミウムのような海洋窒素固定生物は栄養塩の制限に対処しなければならない。先に議論したように、モリブデンは現代の海洋で容易に入手可能であるが、鉄は主要な制限栄養素であり、一次生産とニトロゲナーゼのような酸化還元酵素に直接影響を与える [45.](図2)。大陸塵の形の鉄の大気沈着は、この生物制限栄養素を外洋に供給することができる(図2)。トリコデスミウムはシデロフォアをコードする遺伝子を持たないため、大気中のダストから鉄を溶解することはできないが、トリコデスミウムのコロニーはダストの捕獲に適した微小環境を提供する代わりに、他のバクテリアのシデロフォアを利用する [46.]。しかし、すべての表層水が実質的な風塵の流入を受けているわけではなく、表層水の約30~40%が高栄養、低クロロフィル(HNLC)地域に分類されている。鉄源がないため、これらのHNLC領域は、多量栄養素が豊富であるにもかかわらず、生産性(すなわち、クロロフィル)が低いという特徴がある [47.,48.,49.]。鉄はまた、他の大・微量栄養素の制限に加えて、亜熱帯ジャ イアに典型的に見られる低栄養・低クロロフィル領域(LNLC) でも生産性を制限する可能性がある [50.]。
図2
図2現代の生物学的窒素固定率の概略図。
キャプションの全文を表示
大きな画像を見るダウンロード 高解像度画像
陸上環境
モリブデンは土壌と地殻において最も希少な生物量子の一つであるため、陸上環境における栄養素と微量栄養素の利用可能性は、現代の海洋環境とは異なっている [36.]。鉄は海洋では重要な制限栄養素であるが、地殻では非常に豊富な金属である。バナジウムも、土壌中ではモリブデンよりもはるかに豊富である [36.](図2)。地上の土壌や地殻のモリブデン欠乏状態を考えると、代替ニトロゲナーゼがBNFに大きく寄与している可能性が高い [36.,51.,52.]。V-ニトロゲナーゼだけで、北方林におけるBNF率の最大50%に寄与していることがわかっている [51.]。しかし、世界のBNFに対する代替ニトロゲナーゼの寄与は、一般的に控えめである [53.]。陸域環境における代替ニトロゲナーゼの影響を評価する場合、陸域のジアゾ栄養細菌は、共生従属栄養細菌と自由生活ジアゾ栄養細菌の2つのグループに分けられることに注意することが重要である。共生従属栄養生物は、自由共生ジアゾ栄養生物よりも農業的意義があるとして歴史的に研究が進められてきたが、代替ニトロゲナーゼは見つかっていない [36.]。自由栄養性ジアゾ栄養細菌が代替ニトロゲナーゼを持つ一方で、陸上の代替ニトロゲナーゼに関する理解は、自由栄養種よりも農業的に重要な共生ジアゾ栄養細菌に焦点が当てられているため、限定的である。
リゾビア(Rhizobia)やフランキア(Frankia)のような従属栄養性通性細菌は、それぞれマメ科植物や放線菌と共生関係を形成している [54.]。これらの共生ジアゾ栄養細菌は、主に低緯度から中緯度の系で見られ、植物宿主にとって生産性の高い環境に対応している [55.]。土壌中の窒素利用可能量が低くなると、共生細菌は植物の根や結節に勧誘される。この細菌は、有機酸栄養素と引き換えに、植物のために窒素固定を開始する [56.]。ジアゾ栄養共生細菌は、窒素が乏しいときには植物にとって有益であるが、共生細菌のために有機炭素を生成するのに必要なエネルギーを考えると、窒素が利用可能なときに共生細菌を支持するのは不利になる [56.]。したがって、共生が不利になる固定窒素濃度の閾値が存在する。窒素の添加は、土壌、腐葉土、植物根粒など様々な基質にわたって、すべてのバイオームでBNFを阻害することがわかった [54.]。
自由生活型の非共生性通性好気性生物は、共生性生物に比べると研究は少ないが、BNFの3分の1に寄与している [55.]。極地環境のように植生がまばらな場合、自由生活性ジアゾ栄養生物は、クリプトガミズム群落に取り込むことで窒素固定に貢献することができる [55.]。これらのクリプトガミック群集は、コケ類、地衣類、真菌類、シアノバクテリア、および/または藻類で構成されている [57.]。自由生活性のジアゾ栄養細菌は、宿主植物が生存できる環境に限定されないため、これらの共生細菌は、寒冷で乾燥した環境におけるBNFにとってより重要であると考えられる [55.]。
世界の窒素固定率
陸域のBNFによって生産される固定窒素の量を定量化することは,その範囲を考えると本質的に困難であるが,最近のボトムアップ・メタ分析では,陸域生態系は年間52~130 Tgの窒素を生産し,世界の中央値は88 Tg N年-1であると推定されている[55.] ハーバー・ボッシュ・プロセスを介した工業的窒素固定(120 Tg N year-1)は、陸上生物群と同量から2倍の固定窒素を生産している [58.,59.]。陸上と工業的窒素固定率はともに,海洋窒素固定率(100-200 Tg N year-1)よりも低い(図2)。
窒素循環の将来
人間活動によって地球の現代の気候が変化しているため,窒素循環のような地球上の生命を維持するために重要な生物地球化学的循環が,大気組成や海洋pHなどの変化にどのように対応するかを予測することが重要である。気候変動の影響には、窒素固定を促進するものもあれば、抑制するものもあると予測されているからである。まず、大気中の二酸化炭素の増加がジアゾ栄養細菌に与える影響について考える。高CO2に反応して光合成が増加するにつれて、固定窒素の需要も陸上と海洋の両方の環境で増加する [60.,61.]。ジアゾ栄養細菌は、pCO2が増加した条件下で繁栄し、窒素固定が29%促進されると推定されるという意見もある [62.]。高pCO2条件下では、炭素濃縮機構(CCM)がダウンレギュレーションされ、エネルギーを窒素固定に向けることができるようになる [62.]。しかしながら、別の研究では、pCO2を2倍にするとCCMのエネルギー利用が30%減少するものの、細胞全体のエネルギー利用には3%しか影響しないと計算されている [63.]。これらの計算に基づけば、窒素固定に対する高CO2刺激の利点は、窒素固定率に対する他の気候変動パラメーターの負の影響を覆すほどには大きくないかもしれない。
海洋への大気中CO2のフラックスの増加は、最終的に海洋酸性化として知られる海洋pHの低下につながる。鉄はpHが低いほど溶けやすくなるが、溶存鉄がさまざまな有機リガンドに取り込まれるため、鉄の生物学的利用能が低下する可能性がある [64.]。HNLCやLNLC地域のような、すでに鉄が著しく制限された環境では、海洋酸性化は窒素固定に深刻な影響を与えるだろう。鉄が制限された環境で生育したトリコデスミウムは、光化学系Iとクロロフィルα合成遺伝子をダウンレギュレートし、これらのプロセスからニトロゲナーゼに鉄を振り向けることができる [65.]。この同じ研究において、鉄の制限と海洋酸性化の条件が組み合わさった結果、Trichodesmium erythraeumの窒素固定率は全体的に低下した。このことは、海洋酸性化が悪化するにつれて、海洋酸性化による鉄溶解度の増加効果は、窒素固定の抑制を覆すには不十分であることを示唆している。最近の予測では、CO2排出が続くと、トリコデスミウムを介した窒素固定は今世紀末までに27%減少するとされている[63]。トリコデスミウムは海洋環境における主要な窒素固定物質であるため、この減少は海洋窒素循環全体に重大な影響を及ぼす可能性が高い。
しかし、地球規模の温度変化は、ジアゾ栄養塩類に対して、その熱的最大値まで刺激的な影響を与えるであろう。熱帯地域では、BNF種の至適温度範囲は温帯地域よりも高く、29℃から37℃の間であるため、温度上昇によりよく反応するであろう [66.,67.]。鉄に制限されたトリコデスミウムの培養における窒素固定率は、22℃から32℃への温度上昇によって308%も刺激された [68.]。温度が22℃から32℃に上昇すると、鉄制限条件下でのCO2固定率も同様に上昇した [68.]。温度の上昇は、これらの培養物の細胞内鉄含有量を減少させ、鉄の制限を緩和する一つの方法を提示している。この同じ研究で、著者らは、2100年までに気温が上昇するとBNFが76%増加すると予測している [68.]。海洋環境では、トリコデスミウム属とクロコスファエラ属はすでに温暖な海域(~30℃)に生息しており、気候変動によって海域の水温がその熱的最大値(36℃)よりも上昇するリスクがある。気候変動の個々のパラメーターの影響を研究する際、BNF率の変化が多様であることを考えると、その結果、生態系と世界の窒素循環がどのような影響を受けるかは不明である。窒素循環は地球上のすべての生命にとって重要な生物地球化学的循環であるため、環境の変化に応じてBNFが示す可能性のある多様な反応を組み込んだ、より複雑な気候モデルを開発することがますます重要になってきている。
宇宙の生命への影響
地球上の生命はニトロゲナーゼの窒素固定活性に依存しているため、これらの酵素の過去と未来の進化を理解することは、「われわれが知っているような生命」の地球外探索に情報を提供する上で極めて重要である。地球上のニトロゲナーゼの制約のもとで、我々は地球外のBNFの探索を、環境中の利用可能な金属補因子とともに、固定可能な窒素源のある惑星または月を探すことから始めるだろう。現在のところ、N2単体では、太陽系外惑星の大気中で分光学的に検出することは非常に困難である。太陽系内では、窒素と既知の代替検出可能なニトロゲナーゼの基質の両方を追跡することによって、BNFの可能性を衛星機器(例えば、イオンまたは質量分析)で評価することができる。例えば、エンケラドスの間欠泉からは、N2、アセチレン、CO2など、ニトロゲナーゼが還元できるいくつかの基質が検出されている[70]。最近のモデルでは、古代の火星には、非常に少量(nM)の固定窒素源を生成することができる、雷に誘発されたN2固定があった可能性が高いことが示されている [71.]。従って、窒素固定が可能な酵素の進化は、初期の火星においても、初期の地球と同様に(これらの微生物が地球上のものと同様の窒素要求量を持っていると仮定すると)有利であることを示している。地球における窒素固定の進化の研究から、BNFが進化する可能性について考慮すべき2つの進化的要因がある。第一に、ニトロゲナーゼが地球の歴史の初期に進化したことは、他の惑星でも同様のプロセスが進化する可能性があることを示唆している。第二に、窒素が基質として豊富にあり、窒素源を固定する必要があるにもかかわらず、地球上の生物は窒素を固定する方法が一つの酵素に限られている。他の場所の生命が、地球上の生命と同じように迅速に窒素固定のための単一のメカニズムを進化させるかどうかは、まだ不明である。
現在の遠隔検出研究とミッションは、生命検出研究に使用される生物学的起源の有機化合物と定義されるバイオマーカーの探索に焦点を当てている。地球上の主要な金属酵素に関与する金属などの無機バイオマーカーの可能性は、ほとんど見過ごされている。窒素固定と窒素循環についてこの総説で論じたように、我々が知っているような生命にとって基本的な地球上の生物地球化学的循環の多くは、様々な金属循環に直接関係している。生物学的窒素固定と工業的窒素固定(すなわちハーバー・ボッシュ)の両方が、窒素を還元するために金属触媒を利用している。したがって、窒素固定触媒における金属の重要性は、すべての生命に普遍的に存在する可能性がある。したがって、ニトロゲナーゼのような酵素を探すには、他の惑星や衛星で金属が利用できるかどうかを知ることが重要である。また、地球外のニトロゲナーゼ様酵素が、その地球化学的環境で利用可能な他の金属補因子や基質を利用する可能性もある。
結論と今後の展望
現存する生体分子や微生物のプロセスを根本的に理解するためには、これらのプロセスが最初に生まれ、形成された歴史的状況を理解する必要がある。過去の出来事の復元は、同位体シグネチャーのような間接的な痕跡を含む、最古の化石記録に大きく依存している。これらの重要なプロセスの多くは、数十億年前に生じたものである。タンパク質の役割、特に私たちの惑星に現存する生命を定義する複雑性のジャンプの根底にあるタンパク質の相互関連的な方法については、利用可能な記録では十分に解明されていない。これには、地球上の生命の歴史において重要な出来事である窒素固定の起源も含まれる。最近の研究によると、窒素固定なしでは複雑な動物は存在し得なかったという [72.]。このように、人類の文明は、数十億年前に起こった特異な生物学的革新に直接起因している。窒素固定が何十億年もの間、多様な生態学的ニッチでどのように発生し、持続し、増殖したかを理解するためには、統合的なアプローチを取る必要がある。ゲノムデータベースで現存する塩基配列の多様性を分析し、進化モデルやインシリコモデルで絶滅したタンパク質の塩基配列の多様性と機能を探り、実験室で過去にさかのぼって再構成されたプロキシ研究を行うことで、地球化学的記録にはほとんど残っていない、失われた地球の分子史の痕跡にアクセスできるようになるかもしれない。今後起こりうる生態学的、気候学的な課題に対する革新的な対応策の武器は、惑星の激変によって加速された過去の革新的な技術によって明らかにされるかもしれない(「未解決の問題」参照)。
未解決の問題
謝辞
この研究は、NASA Interdisciplinary Astrobiology Research Consortium (ICAR) Metal Utilization and Selection across Eons (MUSE)、NASA LIFE Research Coordination Networkのメンバーであり、NASA Science Mission Directorate (No 80NSSC21K0592)およびNational Science Foundation Emerging Frontiers Award (No 1724090)の資金援助を受けている。有益な議論をしてくれた Kacar 研究所と MUSE ICAR のメンバー、図案の作成に協力し、批判的なフィードバックをくれた Morgan Sobol と Amanda Garcia に感謝する。
参考文献
Söderlund R.
スヴェンソン B.H.
地球規模の窒素循環。
Ecol. Bull. 1976; 22: 23-73
記事で見る
グーグル・スカラー
ファルコフスキー P.G.
窒素循環の進化と海洋におけるCO2の生物学的隔離への影響。
Nature. 1997; 387: 272-275
論文で見る
スコープス (961)
クロスリファレンス
グーグル奨学生
Mus F.
et al.
地球生物学的フィードバック、酸素、およびニトロゲナーゼの進化。
フリーラディック。生物学。Med. 2019; 140: 250-259
論文で見る
スコープス (43)
PubMed
クロス
グーグル奨学生
スミス C.
et al.
カーボンフリーのエネルギー展望におけるハーバーボッシュ・アンモニアの現在と将来の役割。
Energy Environ. 2020; 13: 331-344
論文で見る
クロスフィルム
グーグル・スカラー
ビショップ P.E.
ヨーガー R.D.
代替窒素固定システムの遺伝学と分子生物学。
Annu. Rev. Plant Physiol. Plant Mol. 1990; 41: 109-125
論文で見る
スコープス (96)
クロスリファレンス
グーグル奨学生
アンバー A.D.
ノール A.H.
原生代の海洋化学と進化:生物無機ブリッジ?
科学。2002; 297: 1137-1142
論文で見る
スコープス (879)
PubMed
クロス
グーグル奨学生
Aryal B.
他
変化した窒素循環における亜酸化窒素の排出と気候変動への影響。
Environ. Pollut. 2022; 314120272
論文で見る
スコープス (10)
パブコメ
クロス
グーグル奨学生
メイソン R.E.
他
陸上生態系における窒素利用可能量減少の証拠、原因、結果。
サイエンス。2022; 376eabh3767
論文で見る
スコープス (51)
クロスリファレンス
グーグル奨学生
ペイン R.C.
ら.
酸化した微隕石は、新アルケアン期の高 pCO2 または低 pN2 を示唆している。
Proc. Natl. Acad. Science.
論文で見る
スコープス (18)
PubMed
クロス
グーグル奨学生
ソム S.M.
他
27億年前の地球の気圧は現代の半分以下であった。
Nat. Geosci. 2016; 9: 448-451
論文で見る
スコープス (112)
クロスリファレンス
グーグル奨学生
Shimamura K.
ら.
初期地球における隕石衝突による急激な NH3 生成: 第一原理分子動力学シミュレーション.
Sci. Rep. 2016; 6: 38953
論文で見る
スコープス (12)
PubMed
クロス
グーグル奨学生
マテオ・マルティ E.
他
紫外光触媒による生物学的窒素固定:初期大気と生命への影響。
Sci. Rep. 2019; 9: 15311
論文で見る
スコープ (11)
PubMed
クロス
グーグル奨学生
ナバロ-ゴンサレスR.
他
雷による窒素固定の減少による古細菌の窒素危機の可能性。
Nature. 2001; 412: 61-64
記事で見る
スコープス(201)
PubMed
クロス
グーグル奨学生
グラスJ.B.
他
シアノバクテリアと藻類における金属利用と窒素同化の共進化。
Geobiology. 2009; 7: 100-123
論文で見る
(137件)
PubMed
クロス
グーグル奨学生
Stüeken E.
他
3.2Gyrのモリブデンニトロゲナーゼによる生物学的窒素固定の同位体的証拠。
Nature. 2015; 520: 666-669
論文で見る
スコープス (175)
PubMed
クロス
グーグル奨学生
アンバー A.D.
他
偉大な酸化イベントの前に酸素の匂い?
サイエンス。2007; 317: 1903-1906
論文で見る
スコープス (758)
PubMed
クロス
グーグル奨学生
プラナフスキーN.J.
他
大酸化現象の5億年前における酸素光合成の証拠。
Nat. Geosci. 2014; 7: 283-286
論文で見る
スコープス (394)
クロスリファレンス
グーグル奨学生
ジャブウォンスカ J.
タウフィック D.S.
酸素利用酵素の進化は、初期の生物圏の酸素化を示唆している。
Nat. Ecol. Evol. 2021; 5: 442-448
論文で見る
スコパス (51)
PubMed
クロス
グーグル奨学生
ロゼット D.
マンシー S.S.
金属は、我々がそれを知っているように生命に不可欠である。
最前線。Cell Dev. Biol.
論文で見る
スコープス (4)
PubMed
クロス
グーグル奨学生
エヴァンス G.N.
他
太平洋、ファンデフカ海嶺、メインエンデバーフィールドからの熱水噴出流体の時系列における栄養遷移金属。
Earth Planet. 2023; 602117943
論文で見る
Google Scholar
Lyons T.W.
et al.
地球初期の海洋と大気における酸素の増加。
Nature. 2014; 506: 307-315
記事で見る
スコープス (1678)
PubMed
クロス
グーグル奨学生
コリアー R.W.
北東太平洋におけるモリブデン。
Limnol. 1985; 30: 1351-1354
論文で見る
スコープス (270)
クロスリファレンス
グーグル奨学生
ガルシア A.K.
et al.
ニトロゲナーゼの進化史の再構築:祖先のモリブデン-補酵素利用の証拠。
Geobiology. 2020; 18: 394-411
論文で見る
スコープス (36)
PubMed
クロス
グーグル奨学生
ムーア E.K.
et al.
深部時間におけるバナジウムの酸化還元化学と生物学的利用能の進化。
Geobiology. 2020; 18: 127-138
論文で見る
スコープ (19)
PubMed
クロス
グーグル奨学生
レーダー D.
バナジウムは原始生命体の活動において、これまで予想されていたよりも万能な標的なのか?
組織。Biomol. Chem. 2008; 6: 957-964
論文で見る
スコープス (57)
PubMed
クロス
グーグル奨学生
デ・ドゥーヴC.
特異点: 生命の道しるべ。
ケンブリッジ大学出版局、2005年
記事で見る
スコープス (94)
クロスリファレンス
グーグル奨学生
ガルシア A.K.
et al.
ニトロゲナーゼの前身の再構築から、マチュラーゼ様タンパク質が起源であることが示唆された。
Genome Bio. 2022; 14evac031
論文で見る
パブコメ
グーグル奨学生
レイモンド J.
et al.
窒素固定の自然史
Mol. Biol. 2004; 21: 541-554
論文で見る
スコープス (600)
PubMed
クロス
グーグル奨学生
Pi H.-W.
ら.
原核生物における窒素固定の起源と進化。
Mol. Biol. 2022; 39msac181
論文で見る
スコパス(5)
クロスリファレンス
グーグル奨学生
Zerkle A.L.
他。
全微生物ゲノムから推定される経時的な生物地球化学的シグネチャー。
Am. 科学雑誌 2005; 305: 467-502
論文で見る
スコープス (155)
クロスリファレンス
グーグル奨学生
トーマス A.
et al.
生存者バイアスは、再構築された祖先タンパク質の安定性を過大評価する。
bioRxiv. 2022; (Published online November 25, 2022. https://doi.org/10.1101/2022.11.23.517659)
論文で見る
Google Scholar
Schwartz S.L.
et al.
初期のニトロゲナーゼの祖先は、新しい活性部位の多様性を包含していた。
Mol. Biol. Evol. 2022; 39: 1-7
論文で見る
スコープ (3)
クロスリファレンス
グーグル奨学生
ガルシア A.K.
et al.
ニトロゲナーゼの復活と特異な酵素機構の進化。
eLife. 2023; 12e85003
論文で見る
スコープス (1)
クロスリファレンス
グーグル奨学生
パーソンズ C.
他。
地球史における環境遷移を追跡する生命樹を横断する窒素代謝酵素の放射。
Geobiology. 2021; 19: 18-34
論文で見る
スコープ (22)
PubMed
クロス
グーグル奨学生
ハリス D.F.
他
3種のニトロゲナーゼにおけるE4(4H)ヤヌス状態でのN2還元とH2生成の速度論的理解。
Biochemistry. 2018; 57: 5706-5714
論文で見る
スコープス (38)
PubMed
クロス
グーグル奨学生
ベレンジャーJ.P.
他
陸上生態系における代替ニトロゲナーゼによる生物学的窒素固定:総説。
Biogeochemistry. 2020; 149: 53-73
論文で見る
スコープス (64)
クロスリファレンス
グーグル奨学生
フィリッピ M.
他。
紫硫黄細菌は、低モリブデン原生代海洋アナログにおいて、モリブデンニトロゲナーゼを介して窒素を固定する。
Nat. Commun. 2021; 12: 4774
論文で見る
スコパス (13)
パブコメ
クロス
グーグル奨学生
Kaçar B.
他
生体必須元素の進化史は、宇宙における生命探索の指針となる。
ChemBioChem. 2021; 22: 114-119
論文で見る
スコープ (11)
PubMed
クロス
グーグル奨学生
ラムレヒトN.
他。
米国中西部上流に位置する2つのフェルギナス・メロミクティック湖の地球化学的特徴付け。
J. Geophys. Res. Biogeosci. 2018; 123: 3403-3422
論文で見る
スコープス (18)
クロスリファレンス
グーグル奨学生
グルーバー N.
海洋窒素循環: 概要と課題。
アカデミックプレス、2008年
記事で見る
スコープス (307)
クロスリファレンス
グーグル奨学生
Paerl H.
自然水におけるシアノバクテリアの窒素固定パラドックス。
F1000Res.
論文で見る
スコープス (29)
PubMed
クロス
グーグル奨学生
バーグマンB.
他
Trichodesmium-珍しい窒素固定特性を持つ広範な海洋シアノバクテリウム。
FEMS Microbiol. Rev. 2013; 37: 286-302
論文で見る
(171件)
PubMed
クロス
グーグル奨学生
Staal M.
et al.
熱帯海洋における温度によるN2固定シアノバクテリアの排除。
Nature. 2003; 425: 504-507
論文で見る
スコープス (136)
PubMed
クロス
グーグル奨学生
Luo W.
et al.
TrichodesmiumにおけるN2固定は、光合成からの空間的分離を必要としない。
Msystems. 2022; 7e00538-22
論文で見る
スコープ (7)
クロス
グーグル奨学生
ウェイド J.
et al.
進化の原動力としての惑星鉄の時間変化。
Proc. Natl. Acad. Sci. U. S. A. 2021; 118e2109865118
論文で見る
スコープ (13)
クロスリファレンス
グーグル奨学生
Basu S.
et al.
海洋藍藻トリコデスミウムのコロニーは、関連細菌と相互作用して塵から鉄を獲得する。
Commun. Biol.
論文で見る
スコープス (37)
PubMed
クロス
グーグル奨学生
ムーア C.M.
他
海洋栄養塩供給のプロセスとパターン。
Nat. Geosci. 2013; 6: 701-710
論文で見る
スコープス (1340)
クロスリファレンス
グーグル奨学生
ポリヴィウ D.
et al.
世界的に重要なジアゾ栄養細菌トリコデスミウムの鉄源としての砂漠の塵。
Front. Microbiol. 2018; 8: 2683
論文で見る
スコープス (18)
PubMed
クロス
グーグル奨学生
ハッチンス D.A.
カポネ D.G.
海洋窒素循環:新たな展開とグローバルな変化。
Nat. Rev. Microbiol. 2022; 20: 401-414
論文で見る
スコープス (47)
PubMed
クロス
グーグル奨学生
Zhang C.
et al.
富栄養化、LNLCおよびHNLC海洋帯におけるダスト添加による植物プランクトン群集の変動。
Sci. Total Environ. 2019; 669: 282-293
論文で見る
スコープス (28)
PubMed
クロス
グーグル奨学生
Darnajoux R.
et al.
北方林における生態系規模の代替バナジウムニトロゲナーゼ活性のモリブデン閾値。
Proc. Natl. Acad. Sci. U. S. A. 2019; 116: 24682-24688
論文で見る
スコープス (42)
PubMed
クロス
グーグル奨学生
Darnajoux R.
他
北方シアノリケンの代替ニトロゲナーゼによる生物学的窒素固定:モリブデン利用可能性の重要性と現在の生物学的窒素固定推定への影響。
New Phytol. 2017; 213: 680-689
論文で見る
スコープス (47)
PubMed
クロス
グーグル奨学生
ハーウッド C.S.
鉄ニトロゲナーゼとバナジウムニトロゲナーゼ:フェイルセーフ酵素かそれ以上か?
Annu. Rev. Microbiol. 2020; 74: 247-266
論文で見る
スコープス (37)
PubMed
クロス
グーグル奨学生
Zhang X.
et al.
地球規模の窒素循環:窒素収支と動態に重要な酵素、生物、プロセス。
Chem. Rev. 2020; 120: 5308-5351
論文で見る
スコープス (114)
PubMed
クロス
グーグル奨学生
デイヴィス-バーナードT.
フリードリングシュタイン P.
陸域自然生態系における生物学的窒素固定の世界的分布。
Glob. Biogeochem. Cycles. 2020; 34e2019GB006387
論文で見る
スコープス (60)
クロス
グーグル奨学生
Pfau T.
et al.
代謝モデリングによって解明された共生窒素固定における絡み合った代謝。
Sci. Rep. 2018; 8: 12504
論文で見る
スコパス (33)
PubMed
クロス
グーグル奨学生
Durán J.
et al.
極地の陸上生態系における土壌の属性と機能を決定する隠蔽率。
科学。Total Environ. 2021; 762143169
論文で見る
スコープス (9)
パブコメ
クロス
グーグル奨学生
ファウラーD.
他
21世紀の地球規模の窒素循環。
Philos. トランス。R. Soci. B Biol. 2013年; 36820130164
論文で見る
グーグル奨学生
ヴィトセック P.M.
et al.
生物学的窒素固定:陸上生態系における速度、パターンと生態学的制御。
Philos. Trans. R. Soci. B Biol. 2013年; 36820130119
論文で見る
グーグル奨学生
レヴィタン O.
et al.
海洋藍藻トリコデスミウム(Trichodesmium)の窒素固定と成長を促進するCO2濃度上昇。
Glob. Chang. 生物 2007; 13: 531-538
論文で見る
スコープス (210)
クロスリファレンス
グーグル奨学生
ロジャース A.
et al.
二酸化炭素濃度の上昇は、マメ科植物における窒素固定の利益を増幅するか?
Plant Physiol: 1009-1016
論文で見る
スコープス (209)
PubMed
クロス
グーグル奨学生
ワニッケ N.
他
海洋酸性化に対する海洋窒素循環の反応。
Glob. Chang. Biol. 2018; 24: 5031-5043
論文で見る
スコープス (35)
PubMed
クロス
グーグル奨学生
Luo Y.W.
et al.
世界的に重要な窒素固定生物トリコデスミウムの海洋酸性化に対する応答は、ニトロゲナーゼ効率の低下が支配的である。
Nat. Commun. 2019; 10: 1521
論文で見る
スコープス (30)
PubMed
クロス
グーグル奨学生
Shi D.
et al.
海洋酸性化が海洋植物プランクトンの鉄利用性に及ぼす影響。
Science. 2010; 327: 676-679
論文で見る
スコープス (282)
PubMed
クロス
グーグル奨学生
Zhang F.
et al.
海洋性ジアゾ栄養細菌トリコデスミウムの鉄過剰および鉄制限条件下における海洋酸性化に対するプロテオーム応答。
Photosynth. Res. 2019; 142: 17-34
論文で見る
スコープス (9)
PubMed
クロス
グーグル奨学生
ビトネロビッチT.A.
他
樹木の窒素固定は樹種や生育温度によって温度感受性が異なる。
Nat. Plants. 2022; 8: 209-216
論文で見る
スコパス (12)
PubMed
クロス
グーグル奨学生
ワン Y.P.
ホルトンB.Z.
窒素固定の気候転換点。
Nat. Plants. 2022; 8: 196-197
論文で見る
スコパス(2)
PubMed
クロス
グーグル奨学生
Jiang H.B.
et al.
海洋窒素固定における鉄の制限を緩和する海洋温暖化。
Nat. 気候。Change. 2018; 8: 709-712
論文で見る
スコープス(48)
クロス
グーグル奨学生
シュウィーターマン E.W.
et al.
衝突対を用いた惑星大気中のN2存在量の検出と制約。
ApJ. 2015; 810: 57
論文で見る
スコープス (64)
クロスリファレンス
グーグル奨学生
マトソン D.L.
et al.
エンケラドスのプルーム: 高温内部の組成学的証拠。
Icarus. 2007; 187: 569-573
記事で見る
スコープス (121)
クロス
グーグル奨学生
アダムス D.
et al.
初期火星における窒素固定。
Astrobiology. 2021; 21: 968-980
論文で見る
スコープス (7)
PubMed
クロス
グーグル奨学生
アレンJ.F.
他
ニトロゲナーゼの阻害は、地球の原生代の大気の酸素化を制限した。
トレンド植物科学 2019; 24: 1022-1031
論文で見る
スコープス (23)
PubMed
要旨
全文
全文PDF
グーグル・スカラー
キャトリングD.C.
ザーンル K.J.
アーケアの大気
Sci. アドバンス 2020; 6eaax1420
論文で見る
スコープス (210)
クロスリファレンス
グーグル奨学生
齋藤真也.
他
古代海洋の生物無機化学:アルケアン-前生代境界におけるシアノバクテリアの金属要求量と生物地球化学サイクルの共進化?
Inorg. Chim. Acta. 2003; 356: 308-318
論文で見る
スコープス (334)
クロス
グーグル奨学生
ボイド E.S.
他。
モリブデン依存性ニトロゲナーゼの後期メタン生成起源。
Geobiology. 2011; 9: 221-232
論文で見る
(111件)
PubMed
クロス
グーグル奨学生
論文情報
出版履歴
オンライン公開 2023年4月13日
出版段階
インプレス、修正校正
識別
DOI: https://doi.org/10.1016/j.tim.2023.03.011
著作権
© 2023 The Authors. エルゼビア社発行
ユーザーライセンス
Creative Commons Attribution - NonCommercial - NoDerivs (CC BY-NC-ND 4.0)|情報アイコンの再利用方法
サイエンスダイレクト
ScienceDirectでこの記事にアクセスする
図
図1
図1地球地球化学と窒素固定の年表。
図2
図2現代の生物学的窒素固定率の模式図。
関連記事
広告
研究ジャーナル
細胞
癌細胞
細胞化学生物学
細胞ゲノム
細胞宿主と微生物
細胞代謝
細胞レポート
セルレポーツ医学
セルレポートメソッド
セルレポート 物理科学
セルレポート サステイナビリティ
細胞幹細胞
細胞システム
化学
化学触媒
カレントバイオロジー
発生細胞
デバイス
ヘリオン
免疫
アイサイエンス
ジュール
マター
医学
分子細胞
ニューロン
一つの地球
パターン
STARプロトコル
構造
トレンドレビュージャーナル
生化学
バイオテクノロジー
癌
細胞生物学
化学
認知科学
生態学・進化学
内分泌学・代謝学
遺伝学
免疫学
微生物学
分子医学
神経科学
寄生虫学
薬理学
植物科学
パートナージャーナル
AJHG
生物物理ジャーナル
生物物理学レポート
HGGアドバンス
分子植物
分子治療ファミリー
ネクサス
植物通信
幹細胞レポート
イノベーション
コレクション
ベスト・オブ・セルプレス
セルプレスレビュー
セルプレスセレクション
コンソーシアムハブ
Nucleusコレクション
スナップショット・アーカイブ
ジャーナルを超えて
細胞キャリアネットワーク
細胞シンポジウム
ラボリンク
ウェビナー
論文を進化させる
図360
複数ジャーナル投稿
スニークピーク
STARメソッド
社会における科学
細胞写真ショー
セルプレスポッドキャスト
セルプレスビデオ
ぬりえ&コミック
リサーチ・アーク
コネクト
セルプレスについて
採用情報
お問い合わせ
ヘルプ&サポート
ニュースルーム
出版アラート
アクセス
購読申し込み
今すぐ読む
司書に薦める
インフォメーション
広告主の皆様へ
リクルーターの方へ
図書館員の方へ
利用規約
プライバシーポリシー
アクセシビリティ
本サイトのコンテンツは、あらゆる科学分野の医療従事者および研究者を対象としています。
当サイトでは、サービスの提供・向上やコンテンツのカスタマイズのためにクッキーを使用しています。クッキーの設定を更新するには、このサイトのクッキー設定をご覧ください。
このサイトのすべてのコンテンツ: 著作権 © 2023 Elsevier Inc.
テキストマイニング、データマイニング、AIトレーニング、および同様の技術に関するものも含め、すべての権利はエルゼビア社に帰属します。
すべてのオープンアクセスコンテンツには、クリエイティブ・コモンズのライセンス条件が適用されます。
プライバシーポリシー 利用規約 アクセシビリティ ヘルプ&サポート お問い合わせ
RELX