
基本的な重原子類における神経管の進化的起源

メインコンテンツへスキップ
感動を与える科学
セルプレスジャーナル
ログイン
レビュー|33巻8号、PR319-R331、2023年4月24日発行
全号ダウンロード
基本的な重原子類における神経管の進化的起源


イゴール・アダメーコ
オープンアクセスDOI:https://doi.org/10.1016/j.cub.2023.03.045
PlumX メトリクス
概要
ヒトを含む脊索動物の中枢神経系(CNS)は、脳脊髄液を含む繊毛状の壁を持つ中空の管として発達する。しかし、地球上に生息するほとんどの動物は、このような設計ではなく、神経節と呼ばれる上皮のない神経細胞の凝縮体から中枢脳を構築しており、上皮のある管や液体を含む空洞の形跡はない。動物界では、上皮のないガングリオン型の神経系が主流であるため、チューブ型中枢神経系の進化的起源は謎のままである。ここでは、脊索動物の神経管の起源、組織学、解剖学に関する潜在的な相同性とシナリオを理解するために関連する最近の発見について論じる。例えば、放射状グリアの存在、層状構造、上皮の保持、折りたたみによる形態形成、液体で満たされた内腔の形成などである。最近の発見は、中枢神経系の管状上皮構造を説明する仮説的な進化シナリオの新しい見方を刺激するものである。その一つは、初期の神経管は、液体を含む内部空洞によって促進された方向性のある嗅覚の改良の鍵であったとする考えである。その後、神経管の嗅覚部分が分離し、脊椎動物では独立した嗅覚系と後方管状の中枢神経系が形成されるようになった。別の仮説によると、厚い基底上皮の神経索は新口動物祖先にさらなる生体力学的支持を与え、後に基底上皮の神経索を液体で満たされた管にすることで改善された、水圧骨格と考えられる。
本文
はじめに
脊椎動物の神経科学、特に生物医学の分野では、脊椎動物の脳と脊髄の研究が中心で、基本的な神経管の設計図に従って構築されています
1
. このアプローチは、ヒトの中枢神経系(CNS)を理解するために多大な努力が払われてきたため、人間中心の視点に大きく影響されてきた。その結果、脊椎動物を扱う多くの神経生物学者は、管状CNSのユニークな解剖学と組織学に慣れてしまった。同時に、動物界の大部分の神経系は、異なる発達を遂げており、神経構造が液体の貯蔵庫として機能することとは全く関係がない。
2
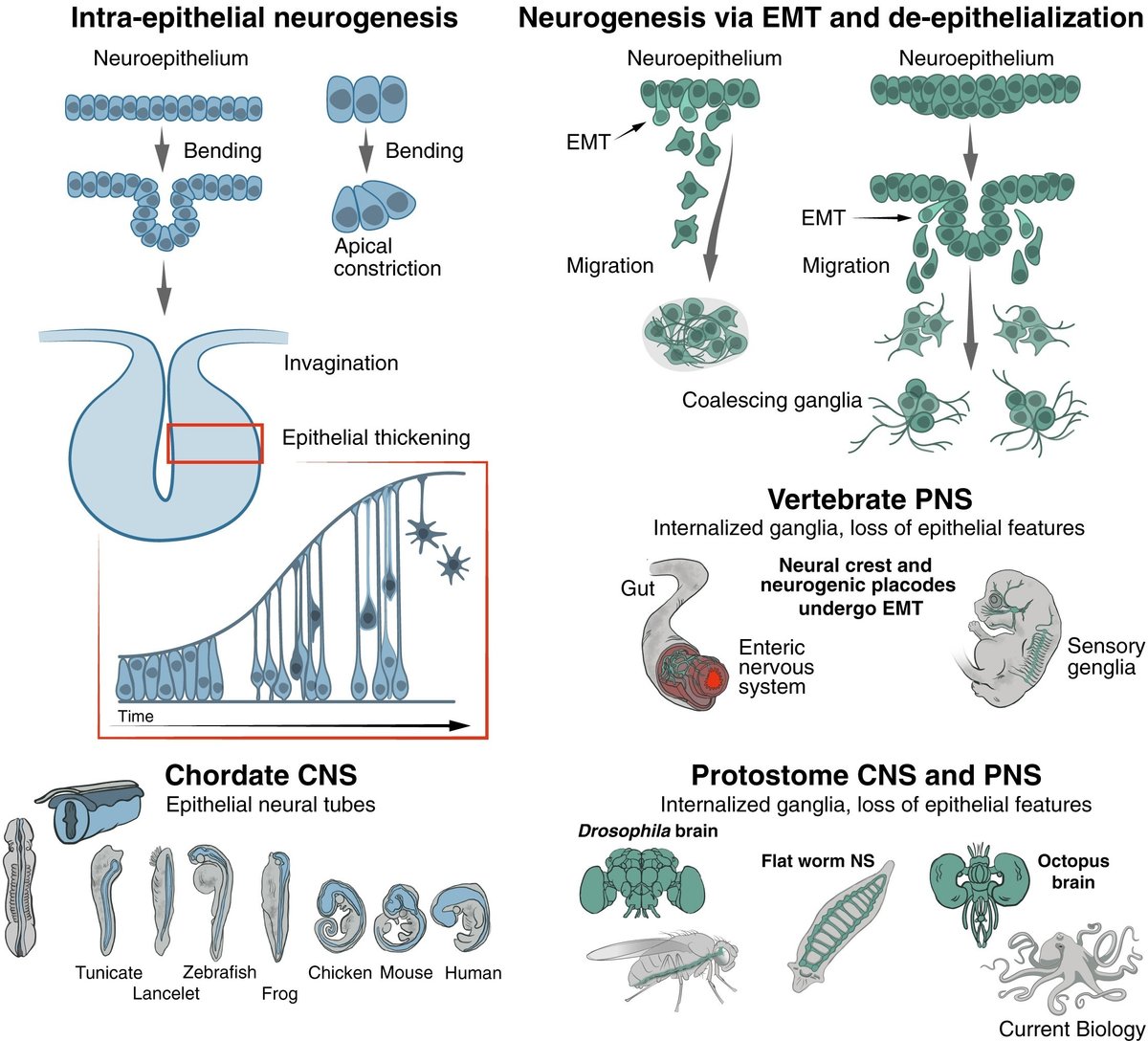
. このような観点から、神経管は、原生生物モデルを扱う神経科学者や進化生物学者の幅広いコミュニティにとって謎めいた存在に見えます(図1)。
図1神経系の形態形成の2つのタイプ。
キャプションの全文を表示する
大きな画像を見る
高解像度画像ダウンロード
脊索動物のチューブ型あるいはリザーバー型の中枢神経系は、ニューロン、グリア、その他の細胞タイプで構成された厚い壁を持つ縦長の血管を表しています。チューブの真ん中にある水路は、上衣細胞で覆われた繊毛状の表面を持ち、液体の流れを積極的に駆動している。脊椎動物では、繊毛で覆われた水路の前部が拡大し、相互に連結した心室が形成されます。この脳室の壁が、発達中の脳と目の感覚器官を構成している。
3
. これまでのところ、神経管の進化に焦点を当てた研究では、神経管のような構造がどこから来たのかという疑問はほとんど無視されていた
4
. しかし、この疑問に触れる前に、上皮化したチューブ型と非上皮化したガングリオン型の神経構造の発達と概念の違いについて議論することが重要である(図1)。
神経節は、神経細胞のソーマ、神経突起、常在グリア細胞、補助細胞(例えば、結合組織カプセルの構築)が凝縮したもので、間葉系組織のコアに浸かっていると定義することができる。神経節の発生は、神経芽細胞(あるいは神経堤のようなブロードタイプの神経・グリア前駆細胞)が神経原性上皮から上皮間葉転換(EMT)を経て剥離することから始まる。前駆細胞は上皮の特徴を失い、間葉系組織に代表される基底膜下の空間に移動し、そこで互いに接着し、狭い細胞集塊の中で神経とグリアの細胞タイプの異質性を発達させます。神経外胚葉からの剥離後、移動する神経原基はEMTにより元の上皮の特徴を失うため、神経節や神経節鎖の発達過程において、上皮組織は保持されず再現もされない
5
.
管状CNSの場合、折りたたみ式神経原性上皮に由来する管壁が
6
上皮の特徴を保持し、神経細胞、グリア細胞、その他の細胞種を宿し、これらの無傷の太い管壁の内部で発達する。対応する上皮の特徴は、堆積した細胞タイプの規則的な層、アピコ基底方向に配向したグリア細胞、基底(または類似)膜、および上皮の特徴が消えることなく、むしろ形態形成的に変形する発生動態を含む
6
. この場合、神経発生上皮は個々の非上皮性移動細胞に分散することなく、上皮の集団行動を維持し、上皮内神経発生に転化する
7
,
8
. したがって、非上皮化したガングリオンに比べて
5
,
9
,
10
,
11
上皮化した神経管の発生は、上皮の形態形成に続いて上皮内神経新生が起こるという、明確で明確なプロセスである(図1)。上皮内神経新生とEMTを介した神経新生との違いは膨大であり、複雑で多因子にわたる進化的に保存された多くのプロセス、例えばEMT
9
方向転換
12
特異的な接着、上皮の折りたたみまたは滑走の調整
7
.
脊索動物の中枢神経系(脳室を持つ脳を含む)は上皮管として組織されているが、脊索動物には神経節的なデザインを持つ非上皮化部分も発達しており、例えば感覚神経系や自律神経系の神経節が含まれている
13
,
14
,
15
(図1)。脊椎動物の中枢神経系は、その発生の全期間にわたって上皮組織に依存しているのとは異なり
6
,
7
これらの神経節は、神経堤や神経原性上皮プラコードに由来する移動性前駆細胞から派生している
16
. これらの前駆細胞は、胚の最終位置に到達するまでに必ずEMTを受け、末梢神経系の体性感覚、交感神経、副交感神経、腸管(腸を支配する)部分を生じさせる。
9
. 興味深いことに、脊椎動物の腸神経系の非上皮化した神経節のニューロン量は、チューブ型の上皮化した脊髄のニューロン量と同程度であった
17
,
18
.
さらに、上皮のない神経節系は、原始・新生動物の種類によってむしろ一般的であり、その中心化の度合いも大きく異なる
5
. 刺胞動物は拡散性の神経網の中で中程度の集中化を示すが、他の動物、例えば節足動物は、脳を含む定型的で比較的大きな非上皮化神経節を示す
19
. さらに、タコの多発性神経節と非上皮性脳は5億以上の神経細胞でできており、これは小型の霊長類の脳に匹敵します
20
. 頭足類の脳の各葉は神経節組織の特徴を保持しているが、脊椎動物の脳の層状構造に類似したいくつかの機能層が出現している。
21
. このように、神経節型やチューブ型のデザインとは無関係に、進化は高度に中枢化した神経系を形成することに成功した。
チューブ型とガングリオン型の神経系は、進化的に保存された地域的な特徴を持ち、外胚葉の起源が一致している。例えば、Six3/6、Otx、Pax2/5/8、Gbx、Hox1遺伝子は新獣類と原獣類の神経構造で同様の領域発現を示している。
22
. 別の例では、環形動物Platynereis dulmeriiのNkとPax遺伝子の発現によって輪郭を描かれる縦方向の神経外胚葉ドメインは、胚性脊椎動物の神経管における特定の縦方向の発達ドメインと対応している。従って、神経系の典型的な縦方向のパターニングは、仮に最後の共通二足動物の祖先に既に存在していた可能性がある。
23
. 最後に、刺胞動物のNematostella vectensisで確認された前後軸のパターン化プログラムは、脊索動物のチューブ型脳領域化を生み出すために、共用して修正されたかもしれない。
4
.
このように、上皮化したチューブ型神経系は、原始生物の神経節型神経系との軸方向パターンや細胞の同一性に基づく類似性から支持されるように、デノボで生じたものではないのだが
4
しかし、その起源は謎に包まれたままである。神経節の集中・拡大による集中型ではなく、チューブ型神経系の進化的起源を促す仮説的な条件や遷移は何だろうか?この問いに答えるために、脊索動物以外の様々なグループの神経系の組織について見てみよう。
棘皮動物の中枢神経系と脊索動物の神経管の相同性
棘皮動物は、半索動物や脊索動物のほかに、羽毛虫やウミユリ(クリノイド)、ヒトデ(アステロイド)、脆性星(オフィユロイド)、ホロ虫、ウニ(エキノイド)など、主要な重母体クレードを代表している(図2)。これらの動物の変態期および変態後の神経発達は、毛帯の下にある神経に代表される幼生期の神経系とは独立して進行する。実際、棘皮動物の幼生は、変態時に幼生体の大部分が消滅し、幼生体に巣食う小さな多能性幹細胞群から成体が完全に発生する、いわゆる破局変態を経て成体へと至る。
図2新口動物群間の系統関係(後述
117
.
フルキャプションを表示
大きな画像を見る
高解像度画像ダウンロード
棘皮動物の成体の変態後の神経系は複雑で、神経輪、神経索、末梢神経節、散在神経、感覚器、神経束など解剖学的に異なる構造からなる。主な中枢部は、口と胃を囲む神経輪と、腕や体の側面に入る橈骨神経索に代表される。
24
(図3A-C)。さらに、棘皮動物の中枢神経系は、相互に連結した外膜部と低膜部に細分化されており、これらの部分は、解剖学的に、薄い結合組織で区切られた外側と内側のバンドとして定義されている。このことは、原始動物や新生動物の神経系が外胚葉性であることと一致している。最も初期の発生段階には、外膜リングの初歩と他の外膜構成要素が滑らかな発生順序で形成され、それに続いて基底神経上皮から低神経サブシステムが出現する。どの段階においても、外膜系と低膜系は神経橋でつながっており、少なくともホロスリンの場合、低膜系が外膜索の内面から発生するまでに確立されている可能性がある
24
. 棘皮動物の神経節や口腔神経輪のような神経集中部は、いずれも原生動物や脊索動物の脳と直接相同化することはできない。
図3棘皮動物中枢と半索動物中枢の組織と発達。
キャプションの全文を表示する
大きな画像を見る
高解像度画像ダウンロード
ウミユリ科とアステロイド科の外耳神経系の解剖学的位置は基底表皮性で、神経孔は基底膜に近い表皮の一部であることがわかる
25
,
26
,
27
. 基底上皮配置の場合、残りの非神経上皮との物理的な接続が失われることはない。他の棘皮動物(ホロトロイド、エキノイド、オフィオロイドなど)は、内腔(「エピニューラル・カナル」とも呼ばれる)を囲む表皮下チューブとして組織された神経系を内蔵していることを示す
24
,
28
,
29
,
30
. この状況は半索動物の神経系の配置に似ており、襟鞘と呼ばれる背索の唯一の管腔を持つ部分が上皮の侵襲により内在化し分離する。半索動物の背索の残りの部分と腹神経索全体は、基底上皮のままである。
棘皮動物の変態変態において、神経外胚葉の内在化がどのように起こるかは、現在のところわかっていない。エキノイドやオフィロイドでは、神経外胚葉の肥厚(神経原性プラコード?)を上顎襞が最終的に包み込み、上顎襞は内在化した上顎環や橈骨洞の上で閉じるとする見解がある
25
,
31
,
32
. しかし、E. fraudatrixの外胚葉からリングの初歩が形成されると、中枢神経系の他のすべての領域はこの初歩の延長から発達し、他の外胚葉の源から発達することはない
24
. これらの場合、棘皮動物神経系の中枢部は、上皮組織を保持したまま、無傷の神経上皮の形態形成的変形(侵入、屈曲、生着)を経て形成される(基底上皮性または内在性)、神経節成分の場合は、神経上皮細胞がEMTと移動を経て内在性神経節を形成するのとは異なる
5
,
10
.
成熟した棘皮動物の神経索の組織構造は、半索動物の神経索や、大部分は脊索動物の中枢神経系に似ている。前述のように、いくつかの棘皮動物(エキノイド、オフィロイド、ホロトロイド)では、神経索は中空で、その壁は上衣上皮からなる
25
,
28
,
33
,
34
脊椎動物の脳にある上衣細胞に似ている。脊髄の構造は、神経細胞体に富む層(脊髄や脳の灰白質に似ている)と、神経回路に富む層(脊椎動物の白質に似ている)に分離している。
35
. これに伴い、ホロスリンの神経索を調べたところ、神経化学的特性の異なる複数の縦方向の神経トラックが中枢神経系内に存在することが明らかになりました
36
脊椎動物の上行・下行脊髄路の構成と解剖学的に類似している。
37
.
さらに棘皮動物では、神経索の神経上皮壁は放射状グリア(または同種の細胞)によって横断されており、このグリアは先端と時には基底の接触点を持つ長い細胞である(図3A-C)。棘皮動物の放射状グリア細胞の多くは、先端部のアンカーポイントが1つだけで、長い基底部の突起がある
38
脊椎動物の上衣グリア細胞にやや似ている。さらに脊索動物に似ているのは、保存されているグリア分子マーカーであるCRF、S-100、ライスナー物質(RS)が存在することである。
38
. また、棘皮動物の放射状グリアは、変成神経新生の細胞源であり、再生時の神経新生の主要な源であると考えられる
39
,
40
,
41
. これは、脊椎動物における上衣グリアと放射状グリアの役割に酷似している。このような上皮内神経新生と放射状グリアからの中枢神経再生は、脊椎動物の最後の共通祖先において進化した可能性があるが、主に脊椎動物で研究されてきたことである
39
.
同時に、節足動物のグリア細胞(Drosophila melanogasterで広範に研究されている)。
42
線虫(線虫で探索)。
43
,
44
とプラナリア(Schmidtea mediterraneaなど)。
45
は、機能的な類似性が進化しているにもかかわらず、異なる発生起源と分子特性を示している。
46
. 原生生物グリア細胞の中には、形態や分布の点で脊椎動物の放射状グリア細胞と類似しているものがあるが
47
一部の例外を除き、保存された遺伝子発現シグネチャーの兆候はない。
48
. これらを総合すると、原始生物と脊椎動物のグリアの相同性には重大な疑問がある
48
,
49
一方、棘皮動物の放射状グリア細胞と脊椎動物のグリア細胞の相同性は、形態、発生、組織的位置に加えて、いくつかの保存されたマーカー遺伝子の発現により、より強い支持を得ている
38
.
棘皮動物の神経索における放射状グリアの集団は、分子レベルでは一様ではない。脆性星の橈骨神経索では、橈骨グリア細胞は転写因子Brn1/2/4(Pou3)を差動的に発現しており、これらの転写因子は脊椎動物の神経発生に関わることが知られているので、橈骨グリア細胞の一部の神経発生促進的な役割、あるいは神経発生的プライミングを強調しているのかもしれない
50
. この考え方に沿って、橈骨神経節の神経節腫脹の神経細胞もBrn1/2/4を発現している(Pou3)。
40
. 放射状グリア細胞の別の集団はMycを発現しており、これも神経原性細胞かグリア原性細胞かの判断に関与している可能性がある。
51
脊椎動物の神経管と同じような状態です。
52
. 最後に、NOTCHシグナル経路は、脊椎動物やショウジョウバエの研究から神経新生に関連する役割がよく知られているが、ホロスリンの中枢神経系再生時の増殖と神経新生の促進にも相関があることが明らかになった
53
,
54
. このように、棘皮動物神経索の特定の放射状グリア細胞の再生および一般的な神経発生能は、脊椎動物の脳における神経発生時の放射状グリア細胞および他のグリアの役割に似ている。
どうやら原生生物グリア細胞も神経発生的な機能を持つようだ
55
と、Brn1/2/4(Pou3)とNotch神経原性遺伝子が原生生物神経系でも発現していることがわかった
5
,
56
. このことは、神経原性というグリア細胞が新しいニューロンを生成する能力が、新口動物系と原口動物系が分離する以前に発達していた可能性があることを意味している。このため、棘皮動物と脊索動物の中枢神経系の相同性を確立する手段として、この基準を用いることは困難である。したがって、棘皮動物の放射状グリア細胞と脊椎動物の放射状グリア細胞との類似性がより高いという判断は、潜在的な基礎的つながりをより精査するためのさらなる実験的研究が必要であることを示している。
脊索動物の神経管と棘皮動物の神経索には多くの類似点があるが、棘皮動物の発達した橈骨神経索は数が多く、複数の腕の放射状の幾何学的形状に空間的に関連付けられている
24
. 棘皮動物の橈骨神経索の数と位置は、脊索動物の神経管や半索動物の神経索の位置とは一致しない。棘皮動物の橈骨神経索は多様で複数の配置があることから、背腹やその他の空間的配置に基づく相同性の割り出しは誤解を招く可能性がある。したがって、神経構造全体(口腔神経輪や背側・腹側特定索など)を相同化することは、それらが特定の生物群に由来する多様なものである可能性があり、問題がある。
一方、細胞タイプ、分子プログラム、組織学、発生機構を相同化することは、建設的で有望であり、これらの基準により、異なる神経系における相同な特徴を追跡するためのより良い事例となるであろう。棘皮動物と脊索動物の神経系に共通する性質として、神経上皮の形態形成と上皮内神経新生があることから、すべての重母動物の最後の共通祖先は、基底上皮型または内部化したチューブ型のデザインを持つCNSの上皮化部分をすでに持っていた可能性がある。
半索動物の中枢神経系と脊索動物の神経管との相同性
半索動物には、ミミズ型の腸鰓類(Enteropneusta)とプランクトン食の翼鰓類(Pterobranchia)があり、その形態は非常に多様である(図2)。脊索動物に近縁な重母動物群である腸鰓類半索動物は、中枢神経系の一部としてチューブ型の神経襟帯を発達させます。この神経上皮襟帯は内部化し、内腔を持ち、背側正中線に沿った動物の襟の短い距離だけにかかっている
57
. 成体半索動物中枢神経系の残りの部分には、基底上皮の腹側と背側の神経索があり、これらは周食道路(枝前神経輪とも呼ばれる)と基底上皮神経網によって相互接続されています
2
,
58
. これらの索は、神経線維の組織的な流れ(「白質」)と、その下にある神経細胞とグリア細胞体の層(「灰白質」)で構成されています。基底上皮の背索は、襟足領域に入ると、内化したチューブ型の襟足索に形態変化する。
まず、プチコデロイドにおける無傷の神経外胚葉の侵入は
59
またはSaccoglossus sp.に摂取される。
60
は、襟足が表層上皮から分離することにつながり、脊索動物の一次神経形成のダイナミクスと同様である
57
(図3Dおよび図4)。第二に、Balanoglossus simodensisの折りたたみ神経板の背面およびその他の側面でBmp2/4、Dlx、Pax3/7およびSoxE遺伝子が空間的に分離して発現することは、脊索動物の神経管における背腹パターニングとの類似性を裏付ける。さらに、折りたたみ神経外胚葉のパターニングにおけるヘッジホッグ(HH)の役割が保存されている可能性があることから、発生メカニズムの相同性が支持されるかもしれない。
61
. しかし、このケースにおけるHHの本当のパターニング機能は直接的に示されなかった。第3に、襟足の解剖学的構造から、脊索動物の屋根板、床板、脳室帯に類似したドメインの存在が明らかになった。第四に、液体で満たされた空洞は、脊索動物の神経孔に類似した神経孔で開口している。
62
. 最後に、半索の襟足は、脊椎動物の神経管と同様に上皮の形態形成を経て、上皮内神経新生に至ります。この上皮内神経新生は、脊椎動物の脊髄と同様の解剖学的方法で、神経伝達物質特異性の異なる路を持つ内側偏在灰白質と外側偏在白質を生成し、放射状グリア細胞の存在も確認される(図3D)。
47
,
62
.
図4脊索動物群における一次神経形成と二次神経形成の発達段階。
キャプションの全文を表示する
大きな画像を見る
高解像度画像ダウンロード
Saccoglossus kowalevskiiの発生中の外胚葉と神経系全体の解析により、前神経稜、zona limitans intrathalamica、isthmic organizerなど、脊椎動物の神経管と相同なシグナル伝達センターの存在を確認(Fgf8/17/18、Sfrp1/5、Hh、Wnt1遺伝子の相同な発現を脊椎動物で確認)した
63
. この議論は、襟足と脊索動物の神経管の直接的な相同性を必ずしも支持するものではなく、むしろよりグローバルなパターン形成の側面に基づくものである。
以上のような共通点があるにもかかわらず、相違点もあり、脊索動物と半索動物の共通祖先から分かれた後に、いかに進化的に派生したかが浮き彫りになっている。例えば、半索動物では背側と腹側に2本の神経索があり、同様の構造を持つ周食道路でつながっているが、脊索動物の神経管は背側に集中した1本である。半索動物の背側神経管が脊索動物の神経管と相同であるか、あるいは腹側神経管がそのような相同性を表すかについては、大きな議論がなされている。この議論は重母動物の共通祖先における背腹逆転の問題や、胚発生過程におけるSHH産生組織の位置づけに関連している
61
,
64
,
65
,
66
,
67
,
68
,
69
.
しかし、棘皮動物の橈骨神経索の状況(上記参照)を考えると、これらの特定の半索動物神経索のどれもが、半索動物体内の位置に基づいて脊索動物の神経管と直接相同である可能性があるかもしれない。さらに、半索動物管型襟足索は、他の神経索、凝縮、神経叢を含むはるかに大きな半索動物神経系のほんの一部分を示しているに過ぎず
58
.
同時に、集中的な基底上皮構造、あるいは内部化したチューブ状の神経上皮構造を生み出す発生プログラムは、半索動物と脊索動物の間で相同的で保存されているのかもしれない
62
,
67
,
70
. この概念に沿うように、放射状グリア細胞や白質と灰白質の層を含む半索状チューブ型カラーコードの組織構造は、半索状基底上皮型背側コードや腹側コードの組織構造と似ている。実際、腸管では、内在化したチューブ部分(カラーコード)は、隣接する基底上皮部分に似ており、内在化とチャネルが出現するために「折り畳みが不十分」であるように見える
62
. 背側と腹側のコードの基底上皮構造の形態形成は、文献から明らかではないが、最も妥当なシナリオは、構造全体を上皮層内に維持する侵食または侵入の抑制を示唆するものである
62
,
71
,
72
,
73
,
74
. これは翼足類の神経系の発達に似ており、中足触手の基部にあるカラー内に表皮内ニューロン群が形成される
75
. このグループの神経芽細胞は、上皮層を越えて周囲の間充織に剥離せず、厚くなった上皮プラコードの中に留まります
71
,
75
,
76
,
77
,
78
. 全体として、棘皮動物と同様に、半索動物における継続的な上皮形態形成(神経索と神経管における上皮内神経形成を含む)は、半索動物と脊索動物の最後の共通祖先が中枢神経系の上皮化した部分を有していたかもしれないという考えを支持している。
基底上皮の索と神経管の起源における発生学的および組織学的な革新性
棘皮動物や半索動物では、基底上皮の神経索は内在化せず、上皮層内にとどまる
79
これは、神経原基の剥離が失敗したと解釈される。同様に、神経管は胚発生の過程で内部化するが、上皮構造は維持されるため、同じ主要な発生様式を共有している。
80
. このことは、神経節系の神経細胞が神経発生過程のある時点でEMTにより上皮組織を失ってしまうこととは異なる。例えば、脊椎動物の神経発生プラコードから発生した感覚神経節は、上皮の特徴を保持している痕跡を見せないが、これはEMTと個々の神経芽細胞の遊走により、発生の早い段階で失われる
15
,
81
,
82
,
83
. 神経細胞の神経節への集合は、神経前駆細胞が神経原性上皮から分離し、その下の間充織に進入して神経細胞とグリア細胞のクラスターを作ることで進行する
19
,
82
,
83
,
84
. 神経原細胞のEMTと分離が始まると、その上皮の性質は次第に消えていくが、基底上皮の神経索やチューブの場合、放射状や上衣のグリア細胞には上皮の性質が保持され、対応する神経構造は常に内部化または非内部化の神経原プラコードと類似している
15
.
上皮化した神経索や神経管の進化におけるもう一つの重要な革新は、膨張した上皮構造の内部で神経前駆細胞が拡大することであり、基本的には、放射状グリアの延長繊維に支えられた巨大な神経原性プラコードである。極性化した放射状神経原細胞と上衣神経原細胞の精緻化により、上皮化した神経管の膨張が可能となり、将来の脳の多層構造が形成された。
神経管は、発生過程において上皮構造を保持しているため、上皮の形態形成の古典的な原理(頂部収縮、上皮細胞接触部の再配列、収束伸展など)に従って内部化と折りたたみを行う。
85
. このような上皮の仕組みにより、神経上皮部が側部外胚葉から分離し、浸潤を経て内生管が形成されるのである
80
. もちろん、これだけでなく、両生類で神経管分離前に起こる、非神経外胚葉が神経外胚葉の上を滑走するなどの方法もあり、内部化の方法はこれだけではありません
86
(図4)。上皮の侵入と外胚葉の滑走は、いずれも頂部の収縮、上皮細胞の接着の局所的な変化などを含む上皮細胞のダイナミクスを制御する必要がある
87
,
88
,
89
. このような古代の上皮形態形成の様式が、内生神経管の形成に応用され、成熟した中枢神経系の上皮組織が一般に保持されていることは、重母動物の神経系に保存されている特徴であると考えられる。上皮形態形成と上皮内神経形成の分子的特異性と進化的に保存されたメカニズムが、神経管形成を特別なものにしている
7
,
8
と、神経節系のEMTを介した発生ダイナミクスとは異質なものである。
5
,
9
,
10
,
11
.
進化の過程で、神経節の神経細胞新生の際にEMTが阻害された結果、上皮内神経細胞新生が起こり、子宮内膜が形成されたと考えられる。その後、EMTでブロックされた神経外胚葉が、一般的な上皮の形態形成と協調して折りたたみと侵入を開始し、内生神経管が誕生した可能性がある。上皮組織と上皮内神経新生の保存は、二次神経形成で観察される間葉系神経前駆体から発生的神経新生が始まり、間葉-上皮移行(MET)を経て上皮化した神経管構造として終わる例外例によってさらに裏付けられる(図4)
80
. この二次神経形成は、頭索動物や環形動物では観察されないことから、派生した機能であると考えるのが妥当である。上皮内神経新生の進化を理解することは、脊索動物の神経管の進化的起源を明らかにするために、今後の研究において最も重要なことである。
最後に、上皮化した索状あるいは管状の神経系と、上皮化しない神経節型の神経系が、すべての重母動物群を通じて同じ生物で共存していることに注目することが重要である。例えば、脊椎動物では、神経堤と神経原性プラコードから移動性神経芽細胞が生まれ、間葉系コンパートメントに進入して体性感覚神経系、自律神経系、腸神経系などの末梢神経系の神経節を形成する。
13
,
15
,
81
,
82
,
83
,
90
. 棘皮動物と半索動物では、神経節が合体したびまん性の神経網と、神経索・神経管が集中した神経網を併存している
2
,
91
. 外胚葉由来の神経前駆細胞の間葉系への移動とEMTを含むその発生様式から、脊椎動物の末梢神経節は、神経細胞とグリアが上皮内に形成されるチューブ型中枢神経系ではなく、むしろ原始生物の神経節型中枢神経系に類似している。このことは、ガングリオン型とチューブ型神経系の基本的な前後左右のパターニング分子が深く相同であり、分岐する中心化様式の始まりに先行していることを意味している。
4
,
23
,
91
.
これらの議論を総合すると、神経原基のEMT、個体移動、接着を介した神経節型の中枢化は、すべての重原子類とほとんどの原生生物群に保存されているため、より古く、一方、上皮化した基底上皮型の神経索型あるいはチューブ型の神経系は、重原子類にしか見られないと考えられる。
チューブ型CNSの進化的起源の概念
神経管の進化的な始まりについて考察することは、かなりの量の推測や憶測を伴うが、それを裏付ける実験的、発生的、組織学的証拠が不十分であることが多い。しかしながら、このような考察は、将来の仮説や実験を導く上で貴重なものとなる可能性がある。神経管内腔に液体が存在することは、管状神経構造の進化を説明するもっともらしい進化的シナリオを形成するのに役立つかもしれない。というのも、両生類や一部の棘皮動物のように、外胚葉が重なり合うことでチューブが形成されることはあり得るからだ。
92
,
93
,
94
. したがって、原理的には、平板なプラコード全体を管状にすることなく内包することに問題はないはずである。
さらに、二次神経形成の場合(図4)でも、筒状のデザインは維持される。これは、体が長くなり、体節の数が増えたことを補う進化の工夫だ
80
. 脊椎動物の二次神経形成(これは二次進化的な革新であると思われる)において、後神経管は特異な方法で形成される:上皮侵入を通過する代わりに、神経中胚葉前駆細胞に由来する間葉系細胞は縦長のクラスターに集合し、間葉系から上皮系への変換を経てこの上皮化神経原性クラスターの中心に内腔を生じる
80
. したがって、このチューブは、上皮化した神経系が内部化する際の上皮の形態形成の単なる結果とは解釈できない。チューブは、古代から進化的に保存されてきた上皮内神経発生を伴う複雑な細胞システムである。
このようなチューブの進化の可能性を説明するために、軸索骨格(ノトコード)が進化する前に、初期の神経チューブが主体軸の水力支持に関与していた可能性があると推論する。この見解は、脳脊髄液の圧力が脊椎動物の脳と頭蓋上部の発達の鍵であるという事実によって支持されている
95
,
96
. 神経管の水力学的な役割は重母動物の共通祖先において、管状デザインを維持する主要な理由の一つであると考えられる(図5A、B)。実際、半索動物や棘皮動物は、移動と摂食に水力システムを多用している。半索動物や棘皮動物は、長くて頑丈なノトコードを進化させる代わりに、加圧された水路や水槽の広範なシステムを備えている
97
. このように、脊索動物の神経管は、両生類と脊索動物を統合する原型的な水力学的器官である可能性があります。興味深いことに、進化の後半になると、脊索動物の脳脊髄液は、複数のシグナル伝達と調節の役割を持つようになった。
95
,
98
,
99
図5管状神経系の起源を示唆する仮想シナリオ。
図5管状神経系の起源を示唆する仮説的シナリオ。
キャプションの全文を表示する
大きな画像を見る
高解像度画像ダウンロード
水力で支えるという考え方はもっともだが、神経原細胞がどのようにしてそのような役割を果たすようになったかは明らかではない。一つのシナリオは、厚く密に詰まった上皮性の神経原性プラコードが、周囲の上皮や下層組織と比較して、より優れた機械的抵抗を与えたというものである(図5A)。この議論は、両生類の頭蓋神経原性プラコードの生体力学的データによって強く支持されている
100
このようなプラコードは、生体力学的な手がかりを介して神経堤の移動に影響を与えたと考えられる。このような頑丈なプラコードが厚みを増し、堅い上皮の膨張を維持することで、放射状のグリア細胞の存在を含む、新口動物CNSの組織学的パターンの起源を説明できるかもしれない。このプラコードが管状に変化することで、前後方向の抵抗が増加し、機械的な役割を向上させることができる。
101
そして、一旦管状構造が達成されると、内部の水圧によってバイオメカニクスを改善し続けた(図5B)。
チューブ型中枢神経系の原初的な役割に関するもう一つの考えは、一般的な化学受容、嗅覚、味覚の受容に言及している。基底上皮神経索の解剖学的および系統学的分布(例えば、基部ウミユリ類と半索動物)に基づき、巨大な繊毛基底上皮神経原基が、有茎ウミユリ類にやや似た基部重母動物の摂食溝で栄養素を感知し毛様体活動を制御することによって摂食の制御に関連していたという仮説が成り立つ(図6)。次に、感度と流体力学を向上させるために、このプラコードの一部がさらに侵食して内部化し、内部に繊毛細胞と化学感覚細胞を持つチューブを形成することができた。このチューブ内での分子の拡散により、外界や口腔に露出した開放神経孔から届くさまざまな分子の勾配を受けやすくなり(後述)、こうした嗅覚の指向性も可能になった(図5C、D)。これは、現代の嗅覚器官の動作に似ており、choanaを介して外界に接続されたチューブまたはサックを表している
102
.
図6ウミユリ科動物の鞍部溝内における神経索から神経管への仮説的進化。
キャプションの全文を表示する
大きな画像を見る
高解像度画像ダウンロード
この仮説をさらに裏付けるものとして、前部神経孔は、発達中のホヤの幼虫の口原基と細い鼻下管でつながっているという事実がある
103
. これまで、神経管内腔と発生中の口との接続は、神経管と口腔が十分に確立された後、遅れて行われると考えられていた。しかし、今回得られた新たなデータから、神経管内腔は神経形成期の終わり頃に出現し、物理的には神経管の最前部に相当することが示唆された。また、このような船尾に特有な結果によれば、口は前方の神経孔から生じていることになる。
103
. これに伴い、脊索動物や脊椎動物では、口原基は前神経境界から発生する
103
. また、半索動物である翼足類では、神経外皮は口縁に近接して形成される
71
. 頭索動物では、例えば鼻下管に似た構造は、口腔と神経管内腔の接続の初歩であった可能性がある。この構造の壁から発生する化学感覚集団が嗅覚器官となり、摂食行動を制御するようになったのかもしれない
104
. 全体として、前部神経孔と神経管全般の進化は、嗅覚と味覚(あるいは摂食全般)と関連していたようだ。
この推論はもっともらしく、真剣に検討する価値があるように思えるが、ホヤの「神経孔」については
103
は、両生類における神経孔に相当しないかもしれません。
81
. さらに、両生類の口は体の左側にある二次的な開口部であり、他の脊索動物の口とは相同性が良くないのかもしれない。したがって、鼻下皮質管に似た構造と思われるものは、相同性解釈の観点からは慎重に扱わなければならない
81
.
次に、半索の襟足の機能は、特にその比較的小さなサイズと局所的な局在との関連で、理解されていない
72
. 半索の神経孔が水孔と口開口部に近いことから、チューブ状の半索が何らかの化学感覚機能を持つという仮説が成り立つ。半索神経孔の開口部周辺には、両生類の神経孔周辺のケーリカーピットの仮説と同様に、化学感覚細胞を含む可能性がある。
105
. 水流のあるチューブは、匂い分子がチューブの長さに沿って拡散して勾配を作り、相対的な濃度を感知することができるため、高いダイナミックレンジ(チューブの長さに比例)を持つ嗅覚の優れたシステムとして機能するかもしれません。さらに、チューブの空洞は、外側の体表にある化学感覚受容体に比べて、大きな粒子や引っ掻きから保護されています(図5C、D、図6)。半索動物の嗅覚についてはほとんど知られていないが、襟足は確かに統合された神経化学感覚構造であることが判明すると予想される。
脊椎動物の胚で、嗅覚(というより単一の鼻後葉)プラコードと前神経板の境界の形成を指示する分子シグネチャーをさらに分析すると、発達中の嗅覚系が隣接する神経管形成から分離する重要な進化的ステップが示唆されるかもしれない。このことは、脊椎動物の最も古い嗅覚器官(サイクロストーム類と絶滅した無脊椎動物)は、対になっておらず、正中線に沿って配置されていることを示唆している。
106
,
107
後葉部とともに関節プラコード(鼻後葉部という)として誘導されます。
16
,
107
. また、脊椎動物以外の動物には嗅覚プラコードが存在しないことから、鼻後葉プラコードが前神経板から分離して、化学感覚や神経内分泌機能の向上に特化したという考え方もできるかもしれません
14
,
15
,
81
,
108
,
109
. 第一に、脊椎動物の嗅覚プラコードは、顔面神経堤由来の間充織に侵入し、内部に空洞を持つ上皮系を構築する(概念的には神経孔を持つ神経管に似ている)。この空洞の壁には、神経管の胚発生と同様に、上皮内神経新生時に誕生した嗅覚神経細胞とグリア細胞が存在する
110
,
111
. 他の神経原性プラコードは、いずれも上皮内モードを使ってニューロンを生成していない。一方、他の神経原性プラコードは、EMTと神経芽細胞の移動によってニューロンを生成する
14
,
15
,
81
,
109
.
第二の論拠は、GnRH(ゴナドトロピン放出ホルモン)産生視床下部ニューロンが嗅覚に由来し、対応する神経原性プラコードから広範囲に移動することに基づくものである。GnRHニューロンの神経原基は、プラコードに出現し、嗅覚に由来する鋤鼻軸索に沿って発達中の脳に移動して定着する。
112
. また、最近、人間の非嗅覚的な認知プロセスが、嗅覚に関連する鼻の吸気パターンと同期しているという興味深い発見がなされました。これらの知見に基づき、本研究の著者らは、脊索動物の脳機能全般を支配する、進化的に古い嗅覚に基づく枠組みの存在を提唱した。
113
.
しかし、前下葉プラコードと嗅覚プラコードの起源については、別の見解もあり、それらは前神経境界の非神経外胚葉から進化し、脊椎動物や脊椎動物の祖先が前神経転写因子を採用することによって、何らかの助けを得た可能性が示唆されている。
15
. このような考え方は、脊索動物における嗅覚系と神経管の進化的なつながりを否定するものである。しかし、嗅覚と神経管が早くから結びついていたことを裏付ける強力な論拠がいくつか存在する。嗅覚腔と神経管は、解剖学的につながっており、恐らくは重母動物の最後の共通祖先において、化学感覚と情報処理機能を兼ね備えた単一の管状体であったと考えるのが妥当であろう。その頃、嗅覚と味覚を提供する化学受容を調整するためには、神経管の管内に液体が流れることが不可欠であった。
機能的には、嗅覚の役割を持つ古代の神経管は、神経や内分泌シグナルを利用して、口腔や前腸で摂食や化学感覚に関与する隣接する繊毛上皮を制御できたと考えられる。この考え方に沿って、両生類では、ハッチェック孔は口腔内の繊毛溝で、摂食、増殖、成長に関連するホルモンを分泌する細胞があると示唆されている
114
. Hatschek'sピットは、ウミユリ科の外肛門繊毛溝に似ており、成体でも神経管と内腔に隣接して配置される。同様に、ウミユリ類では、基底上皮の神経索は隣接する摂食両側溝と空間的に並んでおり、その繊毛上皮は
25
,
26
,
27
,
29
このことは、これらの構造間で化学感覚と運動情報の伝達が行われていることを示唆している。仮説として、ウミユリ類の摂食腔溝の上皮には、化学感覚受容体と、ハッチェック孔のホルモン産生細胞仮説に似た神経内分泌細胞が共存している可能性が考えられる。このことから、重生殖動物の黎明期には、化学感覚神経細胞を持つそのような鞍部摂食溝の繊毛上皮の下にある神経柱から、神経管や基底上皮神経索が生まれたのではないかという仮説が成り立つ(Igor Y. Dolmatovの提唱する考えである)。その場合、基底上皮の神経膜を持つ摂食溝の最も中央の部分はさらに侵食して内部化し、残りの部分は内部化した神経管の下にある摂食溝として制御されたままであったかもしれない(図6)。これは、両生類で観察されるハツカネズミの穴のケースと同様である。
この「嗅覚」仮説に対する批判として考えられるのは、両生類をはじめとする重母動物の一次感覚・化学感覚ニューロンは、表皮や表皮下に点在しているという事実に基づくものだ
81
,
108
,
109
,
115
,
116
. やがて、進化によってこれらの神経細胞の前駆細胞が集まり、鼻下葉状体プラコードを含むプラコードフィールドが形成されたのかもしれない。
81
. しかし、化学感応細胞と機械感応細胞はそれぞれ独立して、重母動物の進化の過程で複数回進化した
81
このことは、進化的に新しい神経堤細胞が複雑さを増している脊椎動物において、嗅覚系やその他の化学感覚構造の進化的起源についての説明を制限する可能性があります。
まとめ
近年、異なる重母動物群の中枢神経系は、従来考えられていた以上に互いに類似していることが明らかになってきた。形態学的、組織学的に共通する特徴は、上皮内神経新生が基底上皮の神経索と神経管の形成につながるという、進化的に保存された共通の発生戦略を示唆しています。しかし、上皮内神経新生と神経管形成がどのような進化過程で生じたかは、依然として不明である。そこで、神経管の進化によって嗅覚や味覚が発達したとする「嗅覚」仮説と、神経管によって骨格が改良されたとする「水力」仮説が提案されている。
最後に、進化的に古い神経管が両方の役割を果たす可能性があるため、神経管起源に関する「嗅覚」説と「水圧」説は調和する可能性がある。内腔を外部の液体で満たすことで、一時的な機械的支持を得ることができる一方、再充填の過程で液体の化学組成を細かく感知することができるかもしれない。あるいは、神経管構造の複数の機能に関しては、動物のライフサイクルの段階が関与している可能性もある。例えば、直接発生する子宮内腔動物の幼虫は、神経管が提供する厚い上皮プラコードと水力サポートの恩恵を受ける可能性があるが、実物大の動物は、神経管を主に嗅覚器官として利用する可能性がある。もちろん、これらのシナリオは極めて推測的なものであり、神経管の起源と進化の実際の傾向を解明するためには、将来の実験的裏付けが必要である。このような仮説的シナリオの構築は、神経管型中枢神経系への進化的移行を理解するための仮説駆動型実験研究を刺激し、方向づけることが期待される。
謝辞
Gerhard SchlosserとMarianne Bronnerには、広範な議論を行い、相同性や嗅覚プラコードや鼻下プラコードなどの様々な関連構造の起源に関する見解を提供してもらったことに心から感謝したい。また、脊索動物の基底上皮神経系と神経管が、クリノイドのような鞍部溝の神経成分に由来することを提唱してくれたIgor Y. Dolmatov (A.V. Zhirmunsky National Scientific Center of Marine Biology, Far Eastern Branch, Russian Academy of Sciences) に感謝します。Igor Y. DolmatovとElena Voronezhskayaとの洞察に満ちた議論は、この原稿を生み出すことになった。最後に、この論文のために素晴らしいイラストを描いてくれたOlga KharchenkoとVictoria Chernikovichに感謝する。I.A.は、European Research Council Synergy Grant 856529, EMBO Young Investigator Program, Bertil Hållsten Research Foundation, Swedish Research Council, Cancer Fonden, Knut and Alice Wallenberg Foundation, Austrian Science Fund (FWF), Paradifference Foundationから支援を受けました。
利害関係の宣言
著者は、競合する利害関係はないと宣言している。
参考文献
カンデル・E.
現代神経科学の起源
Annu. Rev. Neurosci. 1982; 5: 299-303
記事で見る
スコープス (8)
クロスリファレンス
Google Scholar
ヘイノル A.
ロウ C.J.
比較のアプローチを取り入れる:神経系の進化を理解する上で、頑健な系統樹と幅広い発生サンプリングはどのような影響を与えるか。
Philos. Trans. R. Soc. Lond. B. Biol. サイ. 2015; 370: 20150045
記事で見る
スコープス (54)
パブコメ
クロスリファレンス
Google Scholar
スタイルズ J.
ジャーニガン T.L.
脳の発達の基礎知識
Neuropsychol. Rev. 2010; 20: 327-348
記事で見る
スコープス (981)
パブコメ
クロスリファレンス
Google Scholar
ファルティン-ゴンザレスD.
ハヴリラックJ.
レイデン M.J.
脳内制御プログラムは中枢神経系の進化に先行している。
bioRxiv. 2021; (Preprint at)https://doi.org/10.1101/2021.12.10.472178
記事で見る
クロスリファレンス
Google Scholar
ハーテンシュタイン V.
ストールヴェルク A.
初期神経形成の進化
Dev. Cell. 2015; 32: 390-407
記事で見る
パブコメ
アブストラクト
全文表示
全文PDF
Google Scholar
新井洋一
タベルナ E.
神経前駆細胞の極性と大脳皮質の発生。
Front. Cell Neurosci. 2017; 11: 384
記事で見る
スコープス (53)
パブコメ
クロスリファレンス
Google Scholar
ニコロプールーE.
ガレアG.L.
ロロ A.
グリーン・N.D.
コップ A.J.
神経管閉鎖:細胞、分子、生体力学的メカニズム。
Development. 2017; 144: 552-566
記事で見る
スコープス (255)
パブコメ
クロスリファレンス
Google Scholar
山口由美子
三浦正治.
脳の形成と閉鎖のしくみ:哺乳類の頭蓋神経管閉鎖のメカニズムへの洞察
Cell Mol. ライフサイエンス. 2013; 70: 3171-3186
記事で見る
スコープス (49)
パブコメ
クロスリファレンス
Google Scholar
ケロスオ・L.
ブローナー・フレイザー M.
癌に悪いことは胚に良いこと:神経堤の発達におけるEMTの重要性。
Semin. Cell Dev. バイオロジー 2012; 23: 320-332
記事で見る
スコープス (105)
パブコメ
クロスリファレンス
Google Scholar
ハーテンシュタイン V.
ウォダルツ A.
ショウジョウバエの初期神経発生。
Wiley Interdiscip. Rev. Dev. Biol. 2013; 2: 823
記事で見る
クロスリファレンス
グーグルシュラー
Zhu Z.
リュウ J.
Yi P.
Tian D.
Chai Y.
Li W.
Ou G.
プロネール遺伝子が線虫の神経芽細胞の非対称分裂と移動を制御する。
FEBS Lett. 2014; 588: 1136-1143
記事で見る
スコープス (0)
パブコメ
クロスリファレンス
グーグルシュラー
マシューズ・H.K.
マーチャントL.
カルモナ=フォンテーヌC.
クリヤマ・S.
ラレイン J.
ホルト M.R.
パーソンズ M.
マヨールR.
生体内における神経堤細胞の方向性移動は、Syndecan-4/Rac1および非共通Wntシグナル/RhoAによって制御される。
Development. 2008; 135: 1771-1780
記事で見る
スコープス (214)
パブコメ
クロスリファレンス
Google Scholar
イームズ B.F.
ムーレマンス・メデイロスD.
アダメイコ I.
進化する神経堤細胞.
CRC Press, Boca Raton2020
記事で見る
クロスリファレンス
Google Scholar
シュロッサーG.
脊椎動物の胎盤の進化的起源:発生学的研究と他の冬虫類との比較から得られた知見。
J. Exp. Zool. B. Mol. Dev. Evol. 2005; 304: 347-399
記事で見る
スコープス (119)
クロスリファレンス
Google Scholar
シュロッサーG.
感覚・神経分泌細胞型の発達: 脊椎動物の頭蓋プラコード、第1巻(進化的細胞生物学).
第1版。CRC Press, Boca Raton2021
記事で見る
クロスリファレンス
Google Scholar
ヨーク J.R.
ユアン T.
マコーリーD.W.
脊椎動物の頭部における神経堤とプラコードの進化的・発達的関連性: 顎のない脊椎動物からの洞察。
フロントフィジオール.2020; 11: 986
記事で見る
スコープス (8)
クロスリファレンス
Google Scholar
ゴールドスタイン A.M.
ホフストラR.M.
バーンズ A.J.
腸に脳を作る:腸管神経系の発達。
Clin. Genet. 2013; 83: 307-316
記事で見る
スコープス (126)
パブコメ
クロスリファレンス
Google Scholar
サッセリV.
パクニスV.
バーンズ A.J.
腸管神経系
Dev. 生物学 2012; 366: 64-73
記事で見る
スコープス (192)
パブコメ
クロスリファレンス
Google Scholar
エッガー B.
チェル J.M.
ブランドA.H.
ハエから神経幹細胞生物学への洞察。
Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2008; 363: 39-56
記事で見る
スコープス (107)
パブコメ
クロスリファレンス
Google Scholar
デリケールA.
スタイフハルスR.
エラゴズA.M.
メースG.E.
セイントイェンス E.
神経前駆細胞とその子孫の同定から、発達中のタコの脳における長距離移動が明らかになった。
eLife. 2021; 10: e69161
記事で見る
スコープス (10)
クロスリファレンス
Google Scholar
シゲノ S.
アンドリュース P.L.R.
ポンテG.
フィオリートG.
頭足類の脳:脊椎動物との比較を容易にするための現在の知見の概要。
フロントフィジオール. 2018; 9: 952
記事で見る
スコープス (63)
パブコメ
クロスリファレンス
Google Scholar
ヴォレセン T.
シェールホルツM.
ロドリゲス・モンヘ S.V.
レドル E.
トッド C.
ワニンガー A.
軟体動物における脳領域化遺伝子の貝殻フィールドパターニングへの協力。
Sci. Rep. 2017; 7: 5486
記事で見る
スコープス (17)
クロスリファレンス
Google Scholar
デネス A.S.
ジェケリーG.
スタインメッツP.R.
レイブルF.
スナイマン H.
プリュドム B.
フェリエ D.E.
バラヴォワンG.
アレントD.
環形動物の神経索の分子構造は、二足動物における神経系の集中化の共通の起源を支持している。
Cell. 2007; 129: 277-288
記事で見る
スコープス (328)
パブコメ
アブストラクト
全文表示
全文PDF
Google Scholar
マシャノフ V.S.
ズエバ O.R.
ハインゼラーT.
アシャワーB.
ドルマトフ I.Y.
ホロスリンの成体神経系の発生起源:棘皮動物の神経発生の謎を解く試み。
Evol. Dev. 2007; 9: 244-256
記事で見る
スコープス (31)
クロスリファレンス
グーグルシュラー
ハイマンL.H.
無脊椎動物 IV.棘皮動物 棘皮動物門.
マグロウヒル社、ニューヨーク1955年
記事で見る
グーグルシュラー
ハインゼラー T.
ヴェルシュ U.
ウミウシ科
にて: Harrison F.W. Chia F.S. 無脊椎動物の顕微鏡解剖学 14. 棘皮動物門。Wiley-Liss, New York1994: 9-148
記事で見る
グーグルシュラー
チア F.S.
Koss R.
アステロイデア(Asteroidea.
にて: 無脊椎動物の顕微鏡解剖学 14.棘皮動物門. Wiley-Liss, New York1994: 169-246
記事で見る
グーグルシュラー
メルケル K.
レザーU.
ウニ(棘皮動物門)の脊柱管および神経索の超微細構造および組織.
Zoomorphology. 1991; 110: 267-279
記事で見る
スコープス (17)
クロスリファレンス
グーグルシュラー
ハインゼラー T.
ヴェルシュ U.
棘皮動物の神経系とその系統的解釈。
in: Roth G. Wullimann M.F. Brain Evolution and Cognition. Wiley-Spektrum, New York2001: 41-75
記事で見る
グーグルシュラー
マシャノフ V.
ズエバO.
ハインゼラーT.
ドルマトフI.
ホロホロ鳥(棘皮動物)の円周神経輪と橈骨神経索の超微細構造。
Zoomorphology. 2006; 125: 27-38
記事で見る
スコープス (43)
クロスリファレンス
グーグルシュラー
ウビッシュ・L.
棘皮動物における骨格の形成と幼体と幼体の対称性.
Zeitschr. Wiss. Zool. 1913; 104: 119-156
記事で見る
Google Scholar
マクブライド E.W.
Echinus esculentusの発生とE. miliarisおよびE. acutusの発生におけるいくつかの点。
Philos. Trans. R Soc. Lond. B Biol. Sci. 1903; 195: 285-330
記事で見る
クロスリファレンス
Google Scholar
スミス A.B.
棘皮動物門の分類.
Paleontology. 1984; 27: 431-459
記事で見る
Google Scholar
バーンM.
オフィユロイデア(Ophiuroidea)。
にて: 無脊椎動物の顕微鏡解剖学 14. 棘皮動物門(Echinodermata). Wiley-Liss, New York1994: 247-344
記事で見る
Google Scholar
トルエックス R.C.
脊髄の機能的神経解剖学:臨床的意義
Clin. 神経外科 1973; 20: 29-55
記事で見る
クロスリファレンス
Google Scholar
ディアス-バルザック C.A.
ラサロ=ペナ M.I.
バスケス=フィゲロアL.D.
ディアス-バルザック R.J.
ガルシア-アララス J.E. (Garcia-Arraras J.E.)
神経解剖学的解析により明らかになったホロスリンの神経系の多様性。
PLoS One. 2016; 11e0151129
記事で見る
スコープス (18)
クロスリファレンス
Google Scholar
ワトソンC.
ハリソン・M.
マウスの全脊髄分節における主要な上行・下行脊髄路の位置:実際と外挿。
Anat. Rec. 2012; 295: 1692-1697
記事で見る
スコープス (26)
クロスリファレンス
Google Scholar
マシャノフ V.S.
ズエバ O.R.
ハインゼラーT.
アシャワーB.
ナウマンW.W.
グロンドーナ J.M.
シフエンテス M.
ガルシア=アララス J.E.
ナマコ(棘皮動物門:Holothuroidea)の中枢神経系は脊索動物のグリア分泌物に対する免疫染色が陽性であった。
Front Zool. 2009; 6: 11
記事で見る
スコープス (37)
クロスリファレンス
Google Scholar
マシャノフ V.S.
ズエバ O.R.
ガルシア・アララス J.E.
棘皮動物の神経再生に重要な役割を果たす放射状グリア細胞。
BMC Biol. 2013; 11: 49
記事で見る
スコープス (47)
パブコメ
クロスリファレンス
Google Scholar
ズエバ O.
クーリーM.
ハインゼラーT.
マシャノワD.
マシャノフV.
脆性星神経系の複雑な単純性。
Front Zool. 2018; 15: 1
記事で見る
スコープス (17)
パブコメ
クロスリファレンス
Google Scholar
マシャノフ V.
ズエバO.
棘皮動物の放射状グリア。
Dev. Neurobiol. 2019; 79: 396-405
記事で見る
Google Scholar
ハーテンシュタイン V.
ショウジョウバエにおけるグリアの形態的多様性と発生。
Glia. 2011; 59: 1237-1252
記事で見る
スコープス (89)
パブコメ
クロスリファレンス
Google Scholar
シャハムS.
線虫の神経系におけるグリアの発生と機能(Caenorhabditis elegans)
Cold Spring Harb. Perspect. バイオル. 2015; 7a020578
記事で見る
スコープス (35)
クロスリファレンス
Google Scholar
ロレンソ R.
オニヅカ M.
デフランスM.
ローラン P.
単細胞RNA配列決定と分子アトラスの組み合わせにより、線虫のニューロンクラスの新しいマーカーが明らかになった。
Nucleic Acids Res. 2020; 48: 7119-7134
記事で見る
パブコメ
Google Scholar
ガルシア=カストロ H.
ソラナ J.
プラナリアのシングルセル・トランスクリプトミクス:新しいツールは、細胞および進化の特徴に関する新しい洞察を可能にする。
Biochem. Soc. Trans. 2022; 50: 1237-1246
記事で見る
スコープス (2)
クロスリファレンス
Google Scholar
ヴェルクラツキー A.
ホー M.S.
パープラV.
神経膠の進化
Adv. Exp. Med. Biol. 2019; 1175: 15-44
記事で見る
スコープス (7)
パブコメ
クロスリファレンス
Google Scholar
ヘルムC.
カールA.
ベッカーズP.
カウル=シュトレヒト S.
ウルブリヒト E.
クルテシス I.
クアルト H.
ハウゼン H.
バルトロメアス T.
ライヒェンバッハ A.
他
両生類における放射状グリア細胞の初期進化。
Proc. Biol. サイ. 2017; 284: 20170743
記事で見る
スコープス (0)
クロスリファレンス
Google Scholar
ロサダ=ペレス M.
グリア:「ただの接着剤」から複雑な神経系における重要なプレーヤーへ:ハエから哺乳類までの比較的な見方。
J. Neurogenet. 2018; 32: 78-91
記事で見る
スコープス (12)
クロスリファレンス
Google Scholar
ヴェルクラツキーA.
アランズ A.M.
シウバ K.
ペコウスカ A.
神経膠の進化
Ann. N. Y. Acad. サイ. 2022; 1518: 120-130
記事で見る
スコープス (2)
クロスリファレンス
Google Scholar
ドミンゲス M.H.
アユーブA.E.
ラキック P.
POU-III転写因子(Brn1、Brn2、Oct6)は大脳皮質上層細胞の神経発生、分子的同一性、移動先に影響を与える。
Cereb. Cortex. 2013; 23: 2632-2643
記事で見る
スコープス (129)
パブコメ
クロスリファレンス
Google Scholar
マシャノフ V.S.
ズエバ O.R.
ガルシア-アララス J.E.
棘皮動物における神経損傷後のプログラム細胞死と放射状グリア脱分化を制御するMyc。
BMC Dev. Biol. 2015; 15: 24
記事で見る
スコープス (27)
クロスリファレンス
Google Scholar
ジーニン N.
アダメイコ I.
ヴィルヘルムM.
フリッツN.
ウフレンP.
エルンフォース P.
ヘンリクソン M.A.
MYCタンパク質は、前駆細胞の分裂様式を制御することで神経細胞の分化を促進する。
EMBO Rep. 2014; 15: 383-391
記事で見る
スコープス (39)
パブコメ
クロスリファレンス
Google Scholar
マシャノフ V.S.
ズエバ O.R.
ガルシア=アララス J.E.
棘皮動物成体神経系における新細胞の不均一な生成。
Front. Neuroanat. 2015; 9: 123
記事で見る
スコープス (16)
パブコメ
クロスリファレンス
Google Scholar
エングラー A.
チャン・アール
テイラー V.
ノッチと神経新生
Adv. Exp. Med. Biol. 2018; 1066: 223-234
記事で見る
スコープス (4)
パブコメ
クロスリファレンス
Google Scholar
オルテガ A.
オリバレス-バヌエロス T.N.
海洋無脊椎動物の神経細胞およびグリア細胞: 最新情報
Front. Neurosci. 2020; 14: 121
記事で見る
スコープス (9)
パブコメ
クロスリファレンス
Google Scholar
ウォレセン T.
マクドゥーガル C.
デグナン B.M.
ワニンガー A.
POU遺伝子は頭足類の中枢神経系の個々の神経節が形成される過程で発現する。
Evodevo. 2014; 5: 41
記事で見る
スコープス (21)
パブコメ
クロスリファレンス
Google Scholar
カウル S.
シュタッハ T.
鎖骨の発生:半索動物Saccoglossus kowalevskiiの神経形成。
J. Morphol. 2010; 271: 1240-1259
記事で見る
スコープス (0)
パブコメ
クロスリファレンス
Google Scholar
ナイト=ジョーンズ E.W.
Saccoglossus cambrensisの神経系について。(Enteropneusta).
Philos. Trans. R. Soc. Lond. B. 1952; 236: 315-354
記事で見る
クロスリファレンス
Google Scholar
モーガン T.H.
Balanoglossusの発達。
J. Morphol. 1894; 9: 1-86
記事で見る
クロスリファレンス
Google Scholar
ベイトソンW.
バラノグロッサス(sp. incert.)の発生における初期段階。
Q. J. Microsc. Sci. 1884; 24: 208-236
記事で見る
Google Scholar
宮本直樹
和田秀樹
半索動物の神経形成と神経管の起源。
Nat. Commun. 2013; 4: 2713
記事で見る
スコープス (40)
パブコメ
クロスリファレンス
グーグルシュラー
ノマクステインスキーM.
ロッティンガーE.
デュフールH.D.
チェットゥーZ.
ロウ・C.J.
マーティンデールM.Q.
ブルネ J.F.
冬虫夏草の神経系は脊索動物より先に中央化した。
Curr. 生物学 2009; 19: 1264-1269
記事で見る
スコープス (0)
パブコメ
アブストラクト
全文表示
全文PDF
グーグルシュラー
パニ A.M.
マラーキー E.E.
アロノヴィッチ J.
アシマコプロス S.
グローブ E.A.
ロウC.J.
脊椎動物の脳信号中枢の古代の重原子類起源。
Nature. 2012; 483: 289-294
記事で見る
スコープス (172)
パブコメ
クロスリファレンス
グーグルシュラー
ホランド L.Z.
基部重原子類の神経系の進化
J. Exp. Biol. 2015; 218: 637-645
記事で見る
スコープス (22)
クロスリファレンス
グーグルシュラー
ゲルハルトJ.
新口動物祖先
J. セルフィジオール 2006; 209: 677-685
記事で見る
スコープス (24)
クロスリファレンス
Google Scholar
ホランド・L.Z.
カルヴァーリョ・J.E.
エスクリヴァ H.
ラウデV.
シューベルト M.
シメルド S.M.
ユー・ジェイケイ
二枚貝の中枢神経系の進化:単一の起源か?
Evodevo. 2013; 4: 27
記事で見る
スコープス (0)
パブコメ
クロスリファレンス
Google Scholar
マーティン=デュラン J.M.
パン・K.
ボルベ A.
レ・H.S.
フル・A.
キャノン・J.T.
ヨンデリウスU.
ヘイノル A.
二枚貝の神経索の収斂進化。
ネイチャー. 2018; 553: 45-50
記事で見る
スコープス (89)
パブコメ
クロスリファレンス
Google Scholar
ゲルハートJ.
ロウ C.
キルシュナー M.
半索動物と脊索動物の起源。
Curr. Opin. Genet. Dev. 2005; 15: 461-467
記事で見る
スコープス (0)
パブコメ
クロスリファレンス
グーグルシュラー
ニールセンC.
脊索動物の中枢神経系の起源-そして脊索動物の起源。
Dev. Genes. Evol. 1999; 209: 198-205
記事で見る
スコープス (93)
パブコメ
クロスリファレンス
Google Scholar
ゲルハートJ.
脊索動物の体軸の反転:代替案はあるのか?
Proc. Natl. Acad. Sci. USA. 2000; 97: 4445-4448
記事で見る
スコープス (70)
パブコメ
クロスリファレンス
Google Scholar
スクリバ,M. (2015). 比較無脊椎動物の発生学アトラス.アルキコエロマタ論』第1巻~第5巻(ミュンヘン:Verlag Dr. Friedrich Pfeil).
記事で見る
Google Scholar
ルペールE.
半索動物と脊索動物をつなぐキーキャラクター:ホモロジーかホモプラシーか?
Can. J. Zool. 2005; 83: 8-23
記事で見る
スコープス (76)
クロスリファレンス
Google Scholar
ラオ・K.
グランダイスプ(Enteropneusta; spengelidae)の発生について.
J. Morphol. 1953; 93: 1-18
記事で見る
スコープス (13)
クロスリファレンス
Google Scholar
ベイトソンW.
バラノグロッサス・コワレフスキー(Balanoglossus kowalevskii)の発達の後期と、エンテロプネウスタ(Enteropneusta)の近縁性に関する示唆。
Quart. J. Microsc. 1885年; 25: 81-122
記事で見る
Google Scholar
シュタッハ T.
グルール A.
カウル-ストレロー S.
Cephalodiscus gracilis (Pterobranchia, Deuterostomia)の中枢神経系と末梢神経系。
Zoomorphology. 2012; 131: 11-24
記事で見る
スコープス (13)
クロスリファレンス
Google Scholar
シェポティエフA.
翼甲類。Anatomische und histologische Untersuchungen über Rhabdopleura normani Allman and Cephalodiscus dodecalophus M'intosh. Zoologische Jarnbucher.
Abteilung fur Anatomie und Ontogenie der Tiere, Jena. 1906; 23: 463-534
記事で見る
Google Scholar
シェポティエフA.
翼竜の研究 2. Teil, Cephalodiscus dodecalophus M'intosh. 1. Abschnitt: セファロディスカスの解剖学。
Zoologische Jarnbucher, Abteilung fur Anatomie und Ontogenie der Tiere, Jena. 1907; 24: 553-608
記事で見る
グーグルシュラー
レーカンパー G.
ヴェルシュ U.
ディリー P.N.
Cephalodiscus gracilis (Pterobranchia, Hemichordata) の神経節の微細構造.
J. J. Comp. Neurol. 1987; 259: 308-315
記事で見る
パブコメ
クロスリファレンス
グーグルシュラー
中野裕之.
中島由紀夫.
雨宮慎一郎
ウミユリMetacrinus rotundusとフェザースターOxycomanthus japonicusの神経系の発達。
Dev. Genes. Evol. 2009; 219: 565-576
記事で見る
スコープス (21)
クロスリファレンス
Google Scholar
ローリー・L.A.
シヴ・H.
脊椎動物の神経形成の戦略とテレオストの神経管形成の再評価。
Mech. Dev. 2004; 121: 1189-1197
記事で見る
スコープス (168)
パブコメ
クロスリファレンス
Google Scholar
シュロッサーG.
感覚・神経分泌細胞型の進化的起源: 脊椎動物の頭蓋プラコード、第2巻(進化的細胞生物学).
第1版。CRC Press, Boca Raton2021
記事で見る
クロスリファレンス
グーグルシュラー
パヴァン・W.J.
レイブルD.W.
神経堤の感覚神経系とメラノサイト系への仕様変更。
Dev. バイオロジー 2012; 366: 55-63
記事で見る
スコープス (50)
パブコメ
クロスリファレンス
Google Scholar
ヤング H.M.
バーグナーA.J.
シンプソンM.J.
マッキューン・S.J.
ハオM.M.
アンダーソン C.R.
榎本 浩二
Colonizing while migrating: How do individual enteric neural crest cells behave?
BMC Biol. 2014; 12: 23
記事で見る
スコープス (55)
クロスリファレンス
Google Scholar
クンツ T.
クラフト K.F.
テックナウ G.M.
アーバッハR.
ショウジョウバエのキノコ体神経芽細胞の起源と分岐した胚系統の生成。
Development. 2012; 139: 2510-2522
記事で見る
スコープス (58)
パブコメ
クロスリファレンス
グーグルシュラー
ジョシ S.D.
デビッドソン L.A.
上皮の形態形成装置とその臓器組立および組織工学への応用の可能性。
Biomech. Model Mechanobiol. 2012; 11: 1109-1121
記事で見る
スコープス (10)
クロスリファレンス
グーグルシュラー
ホランド・N.D.
ホランド・L.Z.
両生類生物学の浮き沈み:その歴史。
Int. J. Dev. Biol. 2017; 61: 575-583
記事で見る
スコープス (6)
クロスリファレンス
グーグルシュラー
セント・ジョンストンD.
サンソン B.
上皮の極性および形態形成。
Curr. Opin. セルバイオロジー 2011; 23: 540-546
記事で見る
スコープス (107)
パブコメ
クロスリファレンス
Google Scholar
鈴木真一
森田浩之.
上野直樹
脊椎動物の神経管閉鎖に寄与する細胞形状の変化の分子機構。
Dev. Growth Differ. 2012; 54: 266-276
記事で見る
スコープス (62)
パブコメ
クロスリファレンス
Google Scholar
ソーヤー J.M.
ハレルJ.R.
シェマーG.
サリバン=ブラウン J.
ロー=ジョンソン M.
ゴールドスタイン B.
アピカルコンストリクション:形態形成を促進する細胞形状の変化。
Dev. バイオロジー 2010; 341: 5-19
記事で見る
スコープス(317)
パブコメ
クロスリファレンス
Google Scholar
ファーラン A.
ディヤチュック V.
カストリティM.E.
カルボ・エンリケ・L.
アブド H.
ハジャブ S.
チョントロッツェア T.
アックラトヴァ N.
ウソスキンD.
カメネフD.
ら。
多能性末梢性グリア細胞が副腎髄質の神経内分泌細胞を生成する。
サイエンス. 2017; 357: イーアール3753
記事で見る
スコープス(177)
パブコメ
クロスリファレンス
グーグルシュラー
ロウ C.J.
ウー・M.
サリック A.
エバンス L.
ランダー E.
シュタンゲ・トマン N.
グルーバー C.E.
ゲルハルトJ.
キルシュナー M.
半索動物における前後方向へのパターン形成と脊索動物の神経系の起源。
Cell. 2003; 113: 853-865
記事で見る
スコープス (378)
パブコメ
アブストラクト
全文表示
全文PDF
グーグルシュラー
ホランド L.Z.
ラウデV.
シューベルト M.
脊索動物アンフォイクス:発生生物学の新たなモデル生物。
Cell Mol. ライフサイエンス 2004; 61: 2290-2308
記事で見る
スコープス (0)
パブコメ
クロスリファレンス
グーグルシュラー
クープ D.
ホランド・L.Z.
脊椎動物の発生機構を解明するための単純なモデルとしての基底脊索動物アンフィオクス。
Birth Defects Res. C Embryo Today. 2008; 84: 175-187
記事で見る
スコープス (0)
パブコメ
クロスリファレンス
グーグルシュラー
ユー・ジェイケイ
ホランド・L.Z.
頭索類(蛞蝓類):脊索動物の性格の進化を理解するためのモデル。
Cold Spring Harb. Protoc. 2009;https://doi.org/10.1101/pdb.prot5288
記事で見る
スコープス (16)
クロスリファレンス
グーグルシュラー
ガトー A.
モロ・J.A.
アロンソ・M.I.
ブエノD.
デ・ラ・マノ A.
マーティン C.
ニワトリ胚の神経上皮の生存、増殖、神経新生を制御する胎生期脳脊髄液。
Anat. Rec. A Discov. Mol. 細胞進化生物学 2005; 284: 475-484
記事で見る
スコープス (0)
パブコメ
クロスリファレンス
グーグルシュラー
辻川 圭一
齋藤和彦
長坂 敦
宮田俊哉
前骨頭における胚脳とその周囲の頭皮の間の発達的に相互依存的なストレッチャー-コンプレッサー関係。
Dev. Dyn. 2022; 251: 1107-1122
記事で見る
スコープス (0)
クロスリファレンス
グーグルシュラー
具同正樹
五芒星棘皮動物の起源に関する進化のシナリオ-形態決定の水力学的原理からの示唆-.
Acta Biotheor. 2005; 53: 191-216
記事で見る
スコープス (5)
クロスリファレンス
グーグルシュラー
フェイム R.M.
レヒティネン M.K.
脳脊髄液システムの出現と発達的役割。
Dev. Cell. 2020; 52: 261-275
記事で見る
スコープス (81)
パブコメ
アブストラクト
全文表示
全文PDF
Google Scholar
マクドナルド A.
ルーB.
カロンM.
カポリッチ=ディヌッチN.
ハットロック D.
ペトレッカ K.
ブールク G.
ストラットン・J.A.
年齢、地域、種を超えた上衣細胞のシングルセル・トランスクリプトミクスにより、上衣細胞の主要な保存機能として繊毛関連と金属イオン調節の役割が明らかになった。
Front. Cell Neurosci. 2021; 15703951
記事で見る
スコープス (9)
クロスリファレンス
Google Scholar
シェラードA.
マヨールR.
生体内で自己生成された硬度勾配に沿った集団的な運動失調。
Nature. 2021; 600: 690-694
記事で見る
スコープス (40)
パブコメ
クロスリファレンス
Google Scholar
ラザルス B.S.
ベラスコ・ホーガン A.
ゴメス-デル-リオ T.
マイヤーズM.A.
ジャスィーク I.
耐衝撃性生物および生物にインスパイアされた材料および構造に関するレビュー。
J. Materials Res. Technol. 2020; 9: 15705-15738
記事で見る
スコープス (0)
クロスリファレンス
Google Scholar
スアレス R.
ガルシア=ゴンザレスD.
デ・カストロ F.
発達と進化における嗅覚系と鋤鼻系の相互影響。
Front. Neuroanat. 2012; 6: 50
記事で見る
スコープス (36)
クロスリファレンス
Google Scholar
ヴィーマン M.T.
ニューマン-スミスE.
エルナシェフ D.
スミス W.C.
ホヤの口は前部神経孔に由来する:脊索動物の進化における口と神経管の関係を再考する。
Dev. 生物学 2010; 344: 138-149
記事で見る
スコープス (44)
パブコメ
クロスリファレンス
Google Scholar
佐藤 剛.
化学感覚モダリティの精緻化と活動の軌跡:脊椎動物の起源に関する新しい視点。
Zoolog. 科学. 2005; 22: 613-626
記事で見る
スコープス (3)
クロスリファレンス
Google Scholar
トジョア L.T.
ヴェルシュ U.
両生類(Branchiostoma lanceolatum)の頭部領域におけるKollikerおよびHatschekの穴と車輪器官に関する電子顕微鏡的観察。
Cell Tissue Res. 1974; 153: 175-187
記事で見る
スコープス (32)
クロスリファレンス
Google Scholar
ジャンヴィエ P.
発生生物学: 鼻に導かれる。
Nature. 2013; 493: 169-170
記事で見る
スコープス (6)
クロスリファレンス
Google Scholar
オイシ Y.
太田 圭吾
倉久 聡
藤本 聡
倉谷 聡
タガメの頭蓋顔面発達と脊椎動物の進化.
Nature. 2013; 493: 175-180
記事で見る
スコープス (102)
クロスリファレンス
Google Scholar
パッテイ C.
シュロッサーG.
シメルド S.M.
脊椎動物の頭蓋底の進化史--I:細胞タイプの進化。
Dev. Biol. 2014; 389: 82-97
記事で見る
スコープス (58)
クロスリファレンス
Google Scholar
シュロッサーG.
パティーC.
シメルド S.M.
脊椎動物の頭蓋底の進化史 II. 外胚葉のパターニングの進化。
Dev. Biol. 2014; 389: 98-119
記事で見る
スコープス (50)
クロスリファレンス
Google Scholar
ビーテスC.L.
川内 聡
クロッカーC.E.
カロフA.L.
嗅覚上皮における神経幹細胞の同定と分子制御。
Exp. 細胞研究 2005; 306: 309-316
記事で見る
スコープス (216)
パブコメ
クロスリファレンス
Google Scholar
チョイ R.
ゴールドスタイン B.J.
嗅覚上皮: 細胞、臨床障害、そして成体幹細胞ニッチからの洞察。
Laryngoscope Investig. Otolaryngol. 2018; 3: 35-42
記事で見る
パブコメ
クロスリファレンス
Google Scholar
チョー・エイチ・ジェイ
シャン Y.
ウィッティントン N.C.
Wray S.
鼻プラコードの発達、GnRH神経細胞の移動、カルマン症候群。
Front. Cell Dev. Biol. 2019; 7: 121
記事で見る
パブコメ
クロスリファレンス
Google Scholar
パールO.
ラヴィア A.
ルビンソンM.
アイゼンA.
ソローカ T.
モル N.
セクンドL.
ソベルN.
ヒトの非嗅覚性認知は吸入と位相が連動している。
Nat. Hum. Behav. 2019; 3: 501-512
記事で見る
スコープス (60)
パブコメ
クロスリファレンス
Google Scholar
窪川紘一
タンドー Y.
ロイ S.
脊索動物の生殖内分泌系の進化.
Integr. Comp. 生物学 2010; 50: 53-62
記事で見る
スコープス (23)
クロスリファレンス
Google Scholar
ラカリ T.C.
両生類の感覚系:祖先の脊索動物の状態を知る窓。
Brain Behav. 進化論 2004; 64: 148-162
記事で見る
スコープス (124)
パブコメ
クロスリファレンス
Google Scholar
ポンブレット G.
シメルド S.M.
脊椎動物の嗅覚系の進化的起源。
オープンバイオロジー2020; 10200330
記事で見る
スコープス (11)
クロスリファレンス
Google Scholar
スワラ B.J.
スミス A.B.
重母動物系統の解読:分子、形態、古生物学の視点から
Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2008; 363: 1557-1568
記事で見る
スコープス(177)
パブコメ
クロスリファレンス
Google Scholar
ロズィノフ S.V.
腕と腕掌: 棘皮動物の集食性付属物の類似点と相違点の形態形成的基礎。
古生物学(Paleontol. J. 2016; 50: 1598-1609
記事で見る
スコープス (4)
クロスリファレンス
Google Scholar
記事情報
アイデンティフィケーション
DOI: https://doi.org/10.1016/j.cub.2023.03.045
著作権について
© 2023 The Author(s). 発行:エルゼビア・インク
ユーザーライセンス
クリエイティブ・コモンズ 表示(CC BY 4.0)|日本経済新聞社
再利用の方法
サイエンスダイレクト
ScienceDirectでこの記事にアクセスする
数値
図1神経系の形態形成の2つのタイプ。
図2新口動物群間の系統関係(後述
117
.
図3棘皮動物中枢と半索動物中枢の組織と発達。
図4脊索動物群における一次神経形成と二次神経形成の発達段階。
図5管状神経系の起源を示唆する仮説的シナリオ。
図6ウミユリ科動物の鞍部溝内で神経索が神経管に進化する仮説。
関連記事
本サイトのコンテンツは、あらゆる分野の医療従事者や研究者を対象としています。
研究雑誌
セル
がん細胞
セルケミカルバイオロジー
セルゲノミクス
細胞宿主と微生物
細胞メタボリズム
セルレポート
セルレポート医学
Cell Reports Methods
セルレポート 物理科学
セルがサステナビリティを報告
細胞幹細胞
セルシステム
ケム
ケム・キャタリシス
カレントバイオロジー
発生細胞
デバイス
ヘリヨン
イミュニティ
アイサイエンス
ジュール
マター
メド
分子細胞
ニューロン
ワンアース
パターン
STARプロトコル
構造
トレンドレビュー ジャーナル
生物化学系
バイオテクノロジー
がん(Cancer
細胞生物学
ケミストリー
コグニティブサイエンス
エコロジー&エボリューション
内分泌・代謝内科
遺伝学
免疫学
微生物学
分子医学
ニューロサイエンス
寄生虫学
薬理科学
植物科学
パートナージャーナル
AJHG
バイオフィジカルジャーナル
バイオフィジカルレポート
HGGアドバンス
モレキュラープラント
分子療法ファミリー
プラントコミュニケーション
ステムセルレポート
ザ・イノベーション
コレクションズ
ベスト・オブ・セルプレス
セルプレスのレビュー
セルプレスセレクション
コンソーシアムハブ
Nucleus Collections
SnapShotアーカイブ
誌面を越えて
セルキャリアネットワーク
セルメンター
細胞シンポジューム
ラボリンクス
ウェビナーズ
奉加帳
コミュニティレビュー
Figure360
スニークピーク
STARメソッド
社会科学
セル画展
セルプレスポッドキャスト
セルプレスの動画
カラーリングとコミック
リサーチアーク
コネクト
セルプレスについて
採用情報
連絡先
ヘルプ&サポート
ニュースルーム
パブリケーションアラート
ACCESS
サブスクライブ
リード・イット・ナウ
ライブラリアンへの推薦
INFORMATION
広告主様向け
採用担当者様へ
ライブラリアン向け
ご利用条件
個人情報保護方針
アクセシビリティ
当社は、サービスの提供や強化、コンテンツのカスタマイズのためにクッキーを使用しています。クッキーの設定を変更するには、本サイトのクッキー設定にアクセスしてください。
Copyright © 2023 Elsevier Inc. 第三者から提供された一部のコンテンツを除く。