No.8 姿勢および歩行制御のための機能的神経解剖学 REVIEW

【タイトル】
Functional Neuroanatomy for Posture and Gait Control

【なぜこの論文を読もうと思ったか】
・歩行はCPGによる自動化された運動だが、高齢者は環境に適応できず転倒したり、脳卒中患者で麻痺症状は軽度でも歩行が困難な患者を経験する。(小脳疾患など)
・歩行にはCPGだけでなく大脳皮質なども深く関与していると考え、まずは大雑把に学んでみようと思ったため。
※読み返しても分かりやすいように、途中で僕の意見も入れてあります。
【姿勢-歩行制御の一般的なスキーマ】
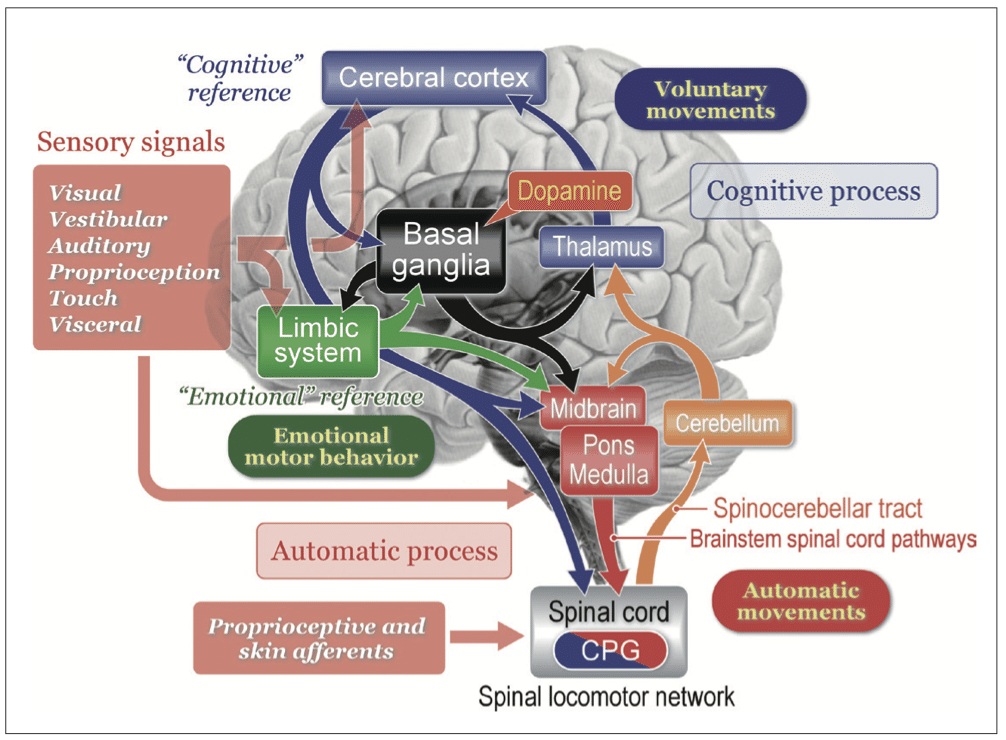
図は運動制御に関係する基本的な、感覚信号からの流れ。
感覚信号は大脳皮質、小脳、脳幹に作用することにより、姿勢の不安定性を検出および修正するために利用される。
動物は、「認知的参照(怖いからどうしよう、トイレに行こうなど)」または「感情的参照(怖い、痛いなど)」のいずれかに応じて動きを開始する。
開始が認知的であるか感情的であるかに関係なく、目標指向の行動は常にバランス調整や筋緊張調節を含む姿勢制御の自動プロセスを伴う。
運動技能を習得したり、なじみのない状況で行動したりするためには、人間は個人外空間における物体の空間的位置特定とともに、自己身体情報の認識に依存する認知姿勢歩行制御を必要とする。
【脳幹および脊髄;姿勢歩行制御のコア構造】
ネコの上丘前縁と乳頭体後縁レベルで除脳が行われると、中脳歩行誘発野(MLR)に電気的または化学的刺激を加えることによって運動を開始する。
MLRに病変が生じた後に、視床歩行誘発野(SLR)に刺激しても、歩行は機械的なものであり、目標指向型に行うことができず、環境へ適応することもできない。
よって、SLRがMLRを超えて脳幹運動経路と接続しており、SLRとMLRの接続は歩行を正常に制御するために重要。
<姿勢と運動の制御における中脳領域の役割>
除脳ネコを使用した実験では、脚橋被蓋核(PPN)のニューロンの活性化は、筋緊張を抑制するが、楔形文字核(CNF)のニューロンの活性化は筋緊張を促進する。
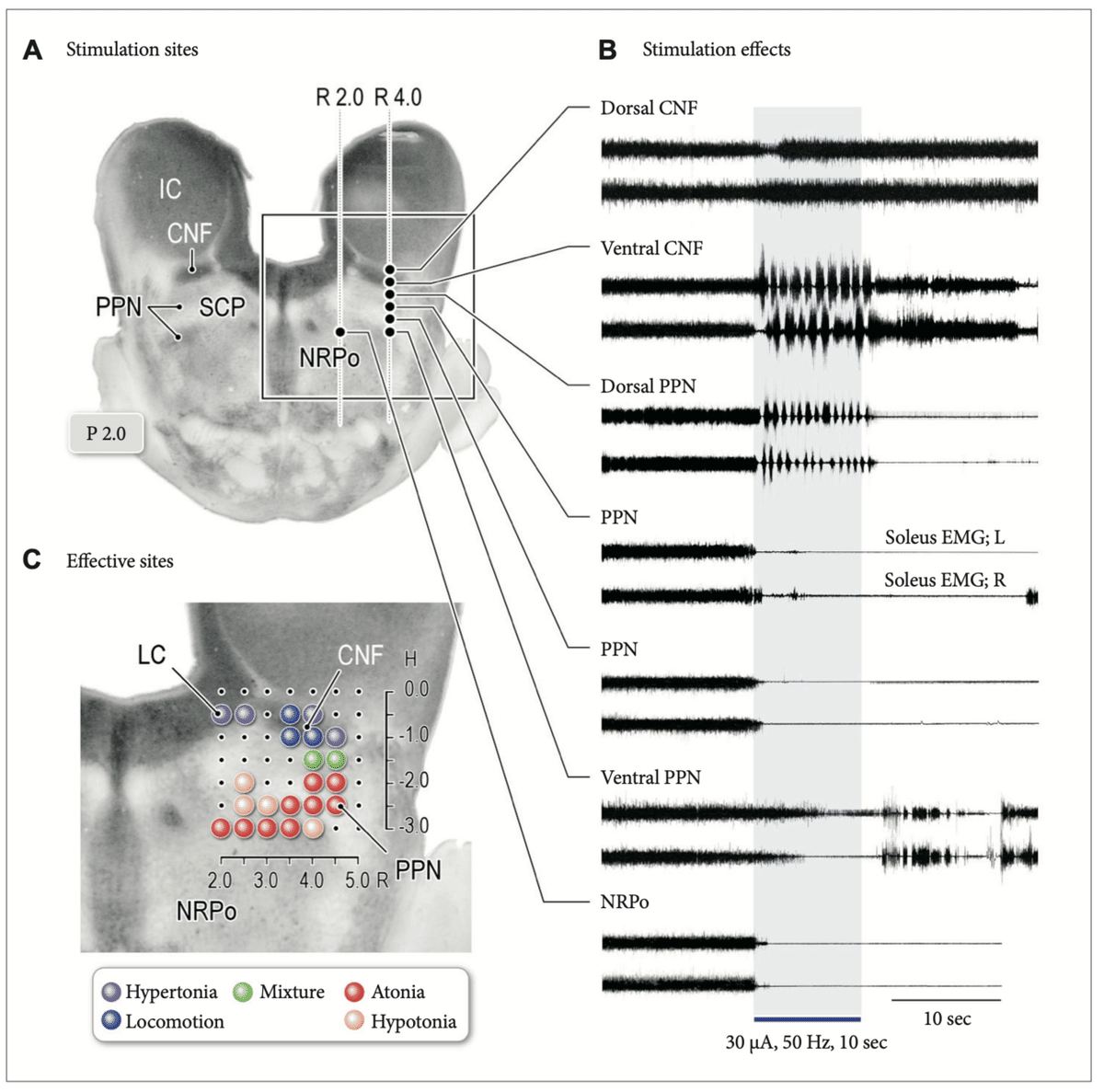
図:徐脳ネコによる中脳刺激の影響
A:右中被蓋の刺激部位
B:Aの各部位に与えられた刺激による筋電図
CNFの背側は筋緊張の増大。腹側CFNと背側PPNへの刺激は運動リズムを誘発したが、背側PPNへの刺激は加えて筋緊張の低下を伴った。
C:トポグラフィー
青斑核(LC)と背外側CNFの刺激は、筋緊張亢進を誘発した(紫;筋緊張増強)。
PPNおよびNRPoの腹外側部分は、筋緊張低下(オレンジ)を誘発した。
運動誘発部位への刺激は、リズミカルな四肢の動きと筋緊張抑制の混合を誘発した(緑)。
CNF:脚橋被蓋核、PPN:脚橋被蓋核、IC:下丘、SCP:上小脳脚
<姿勢制御における網様体の機能的組織化>
網様体脊髄路(RST)が筋肉の緊張レベルの調節に寄与することは一般的に認められている。
姿勢筋緊張の制御に関連して、橋髄質網様体(PMRF)に機能的組織が存在する可能性がある。(図)。
図Aa:PMRFは筋緊張抑制に活動的である。
図Ab:腹内側部分は筋緊張亢進のために活動的。
図Ac:運動関連ニューロンは、運動のステップサイクルに関連するリズミカルな発火を示すニューロンと判断された。

このような組織の存在は、PMRFへの刺激を用いた実験によって裏付けられた。
→上の図のB
図Bの赤:PMRFの背内側部分に適用された刺激は、筋緊張の一般的な抑制をもたらす。
図Bの青:腹内側PMRFに適用された刺激は、筋緊張の一般的な増強を誘発した。
図Bの緑:片側肢の伸展と反対側肢の屈曲を特徴とする被蓋反射または非対称姿勢像は、抑制性領域と興奮性領域の間の領域と、PMRFの外側部分から誘発された。
<運動経路と大脳基底核による制御>
大脳基底核にはサブコンパートメントがある。
①新線条体-背側淡蒼球経路(「背側経路」、図A)
②腹側線条体-腹側淡蒼球経路(「腹側経路」、図B)
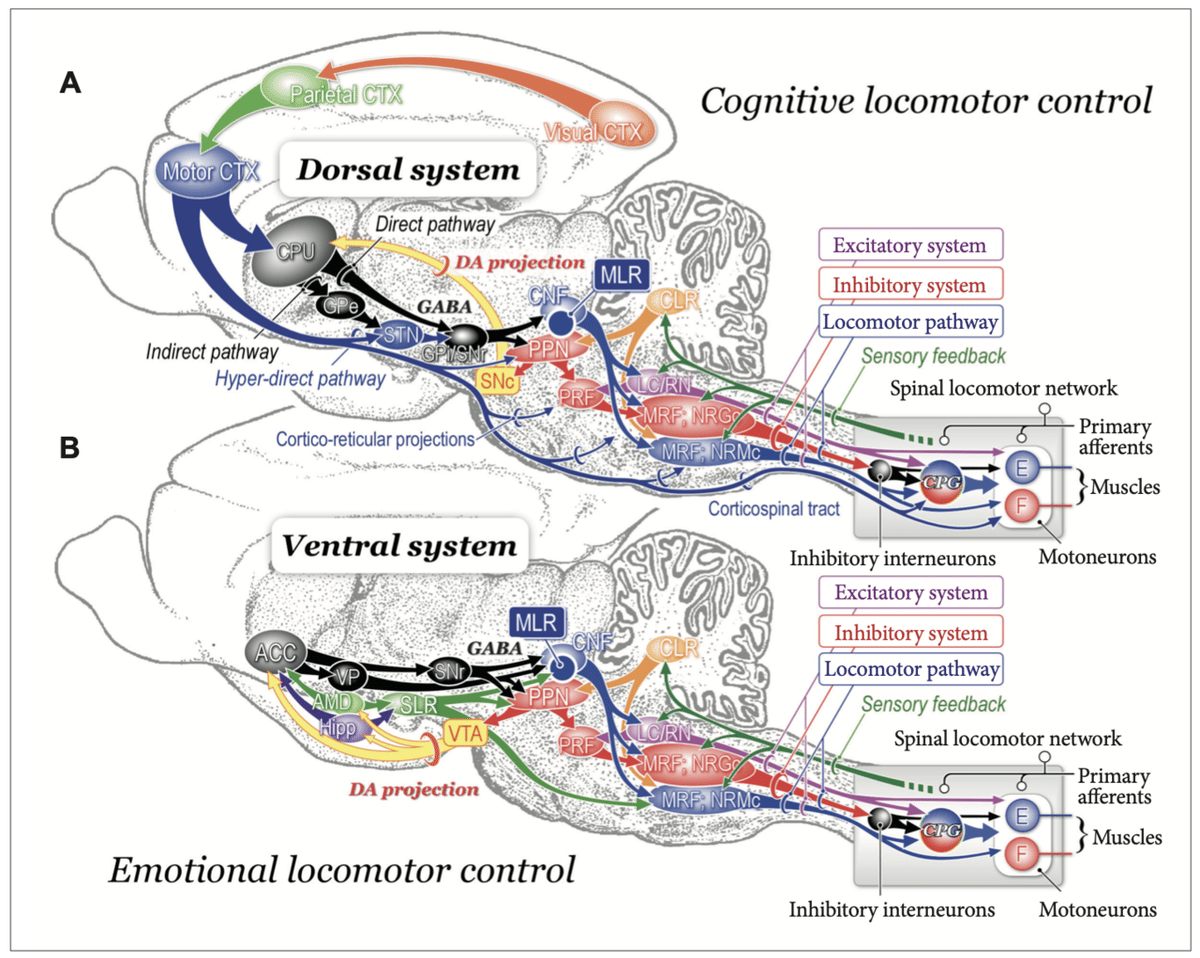
A:認知運動制御のための背側システム
黒質緻密部(SNc)から尾状核-被殻(CPu)へのドーパミン作動性投射は、自発運動の学習に関与している可能性がある。大脳基底核(淡蒼球および黒質網様部の内部セグメント; GPi / SNr)からのGABA作動性出力は、MLR / PPN領域に作用して位置と姿勢を制御する可能性がある。
中脳運動領域(MLR)からの遠心性神経は、興奮性システム、抑制性システム、および運動経路を動員。興奮性システムは、LCと縫線核から生じる。
PPNから生じる抑制性システムは、PRFニューロン、延髄網状脊髄ニューロン、および脊髄抑制性介在ニューロンを順次活性化。抑制性介在ニューロンは、運動ニューロンと介在ニューロンの両方を抑制している可能性がある。
B:感情的な運動制御のための腹側システム
扁桃体(AMD)および海馬(Hipp)からの遠心性神経は側坐核(NAc)に投射。GABA作動性NAcニューロン(ACC)は腹側坐核(VP)に投射。
AMDとHippからの遠心性神経はSLRに作用する。
<姿勢と歩行の脊髄制御>
ストレッチ反射、相互抑制、非相互抑制(または自発的抑制)および屈曲反射に関与する脊髄回路は、姿勢の制御に関与する。
特に、ストレッチ反射とIb抑制は、姿勢の静的制御において主要な役割を果たす。一方、相互Ia抑制と交差伸展反射を含む屈曲反射に関与する介在ニューロンネットワークは、歩行中の左右の脚の交代の伸展-屈曲運動を伴う姿勢像を生成するために重要。
よって、すべての脊髄反射ネットワークの統合は、運動中の筋緊張調節を完全に実行するために不可欠である可能性がある。
<運動中の筋緊張の脊髄制御>
脊髄運動ネットワークの活動は、位相依存的に感覚求心性神経によって調節され、たとえば、股関節の筋肉の固有受容器は、主に立脚相の調節に関与する。
皮膚求心性神経もCPGに強力な影響を及ぼします。皮膚受容体は、障害物を検出し、「つまずき修正反応」などの障害物を回避するために足踏みを調整するために重要。
【姿勢歩行制御の高次規制】
大脳皮質、大脳辺縁系-視床下部構造、大脳基底核などの前脳構造、および小脳は、MLR / PPN領域を介した直接および間接接続を介して網脊髄系に作用することにより、姿勢と歩行を制御する。これらの皮質および皮質下の投射は、動物が文脈に応じて意欲的および感情的な運動行動を表現することを可能にする。
<大脳皮質による制御>
姿勢歩行制御は、空間における身体の向きと動き、および個人外空間における物体の動きの知覚と空間的局在に関する知識に基づいて達成される。このような知識は、小脳、体性感覚、視覚感覚信号の統合によって提供される。
歩行者が障害物に遭遇した場合、各脚は高度に配置する必要がある。
適切な足の配置を達成するために、被験者は各ステップで脚の軌道を変更する必要があるため、このような視覚運動の調整は、四足動物では特に必要です。
動物が障害物を跨ぐときに、視界に障害物は入っていない。しかし動物は安全に跨げる。それには後頭頂葉(上頭頂小葉)が関係。
後頭頂葉は、障害物と障害物との時間空間的関係を認識し覚えておくために必要。短期記憶内の身体スキーマなどの身体情報は、運動皮質で運動プログラムを生成するために利用され、猫が手足の軌道を正確に変更できるようにする。(図B)
歩行中の手足の動きを正確に制御するには、身体の平衡を維持できるように、意図的な行動に先立って姿勢を最適化する必要がある。
したがって、視覚-頭頂-前頭皮質投射(図B)は、姿勢の予測的調整に関与している。

<予測的な姿勢調整>
それでは、運動皮質のどの部分が予測的な姿勢調整のプログラミングに貢献しているのでしょうか?
最も候補となる領域は、SMAとPM。
運動前野(SMA / PM)からPMRFへの豊富な皮質投射が実証されている。
🌟橋髄質網様体(PMRF)は、【姿勢制御における網様体の機能的組織化】で出てきた、背側PMPFが筋緊張抑制に、腹側PMPFが筋緊張促進に関与しているところ。
最近の研究では、SMAが外側PPNに強く接続され、背側PMが内側PPNに接続されていることを示されている。
🌟脚橋被蓋核(PPN)は【姿勢と運動の制御における中脳領域の役割】で出てきた、筋緊張を抑制に関与する部位。
網様体脊髄路(RST)は予測的な姿勢調整を達成する可能性がある(図D)。
おそらく、SMAは、PD患者で損なわれるステップ開始の予測的な姿勢調整に寄与する。
一方、PM / SMAは、正確な脚足の動きのプログラムを運動野(M1)に転送し、M1は皮質脊髄路を介して運動指令を送信する。
特に動物がなじみのない環境に遭遇した場合、正確な歩行制御には頭頂皮質の認知情報が不可欠。
したがって、頭頂から前頭皮質(前頭頭頂ネットワーク)への情報処理の欠陥は、「歩行の凍結」などの予期的な姿勢調整のエラーや歩行の困難を引き起こす可能性がある。
よって、高齢者やアルツハイマー病患者の認知機能の障害は、特により多くの認知課題が必要な場合に転倒するリスクが高くなる。
<垂直性の維持>
脳はどのようにして体の動きと姿勢の垂直性を獲得するのか?
前庭視床投射は両側性であり、主に後部視床を含む。
前庭からのみ入力を受け取る皮質領域はないが、前頭眼野、PM、体性感覚皮質、腹側頭頂内皮質、内側上部側頭野、および頭頂-島前庭皮質(PIVC)に前庭感覚が入力される。
PIVCは、他の前庭関連皮質領域と特に密な関係があり、他の感覚モダリティから情報を受け取る。
現在、視床後部とPIVCの両方が、姿勢垂直性の内部モデルの対象領域。
PIVCは、反対側の前庭神経核への下行性の投射ができるため、前庭皮質系は、姿勢の垂直性の内部モデルに基づいて前庭脊髄系に作用することにより、直立に寄与する可能性があります(図B)。
【小脳による姿勢歩行制御】
小脳による姿勢制御は、感覚求心性神経に大きく依存する。
室頂核(FN)は、脊髄小脳路を介した末梢感覚情報に加えて、遠心性コピー、視覚情報、前庭情報も受け取る。
多感覚機能は、FN歩行の調整などの運動機能に影響を与えることを可能にする「エラー修正メカニズム」を提供する可能性がある。
FNは、高度に統合された身体情報を、脳幹および運動皮質領域の姿勢歩行関連領域に送信する可能性がある。
脳幹への作用
FNから脳幹への出力は、姿勢筋緊張の制御に貢献する。
小脳刺激による筋緊張の促進は、両側の興奮性RSTおよびVSTを活性化によるもの。
小脳刺激による筋緊張の低下は、運動ニューロンへの興奮性の刺激の抑制。
大脳皮質に対する小脳の作用
小脳と大脳皮質の間の相互接続(図C)は、姿勢制御の認知およびプログラミングプロセスに関与している可能性がある。
頭頂葉と小脳の間の相互接続は、空間での体の動きの知覚に関与している可能性がある(図B)。
このような身体情報は、立位時に直立姿勢を維持し、予測的な姿勢調整を実現するために利用できる。
予測姿勢制御は、運動プログラムを構築するために、運動皮質領域(SMA / PM)と小脳(運動ループ)の間の相互接続を伴う可能性があります(図C)。
しかし
小脳疾患の患者では、対照と比較して、予測的姿勢調整の異常はほとんど見られません。小脳は大脳基底核と相互に関連しているため、大脳皮質に加えて大脳基底核が姿勢制御の認知過程の小脳の役割を代償する可能性がある。
大脳基底核が小脳の代償を行う可能性がある。
では、大脳基底核はどのようにして歩行・姿勢を制御しているのか
↓
【パーキンソン病(PD)に関連した大脳基底核による姿勢歩行制御】
PDの姿勢障害は次のメカニズムに起因する。
1)ドーパミン作動性およびコリン作動性システムの障害
2)内部位置を生成することを可能にする統合感覚処理の失敗による認知機能の障害 -構造モデル(ボディスキーマ)
3)運動皮質領域の活動低下による運動プログラミングの失敗、および脳幹の姿勢歩行領域の障害
ドーパミン作動性およびコリン作動性システムの障害
大脳基底核に投射する黒質緻密部(SNc)のドーパミンニューロンの損傷は、大脳基底核からのGABA作動性抑制出力の増加と考えられている。
これは視床、皮質、脳幹を強く抑制している可能性がある。
🌟ドーパミンの不足は、学生時代に習った皮質への影響だけでなくPPNやMLRなどの脳幹へも影響している。
感覚処理と認知の障害
歩行中の対照被験者と比較して、PDの患者の皮質活動は大幅に減少する。
これは統合感覚処理ができず、その結果、内部姿勢モデルと運動プログラミングを妨害する可能性がある。
またコリン作動性PPNの活動の低下が、視床レベルでの多感覚統合を妨げる可能性があることを示唆される。
モータープログラミングの失敗
M1、SMA、PMを含む運動皮質領域は、大脳基底核および小脳と接続しており、大脳基底核からの抑制出力が増加し、自発的運動の実行および運動プログラミングに寄与する運動ループを構成する(図C)。
認知情報処理の低下に加えて、視床皮質の予測は、状況の変化に応じて運動プログラムを生成する能力を低下させる可能性がある。
実際、SMAは、PPNおよびPMRFへの皮質遠心性投射を介してステップ開始の予測的姿勢調整に寄与し、このプロセスはPD患者では深刻に損なわれる。
PDの歩行中に視覚誘導するとPMの背側部分の活動が増加したため、SMA / PMの姿勢歩行プログラムは視覚運動経路の活性化によって促進された。
したがって、PDでの運動プログラミングの失敗は、感覚処理の障害に加えて、運動皮質領域の興奮性の低下が原因である可能性ある。
🌟PD患者は、基底核と接続があるSMAの活動が低下し、予測姿勢制御が困難となり一歩目がなかなか出ない。
視覚情報により、PM背側の活動が上昇したため、視覚情報により予測姿勢制御が利用可能となる。
大脳基底核は、ドーパミンやコリンによって、大脳皮質を活性化し、頭頂葉で感覚の統合、身体図式の生成などを促進するとともに、SMAやPMで運動プログラムを立てられるようにしている。
このようにして、大脳基底核は姿勢・歩行の調節に関わっている。
脳幹への影響はどのようなメカニズム?
↓
中脳の姿勢歩行領域での活動の低下
PPN / MLRを含む中橋被蓋の興奮性の低下は、PDの姿勢歩行障害にも関与している可能性があることを提案される。
黒質緻密部(SNc)のGABA作動性出力経路: SNrの外側部分は、PPNによって誘発される筋緊張抑制をブロックするが、SNrの内側部分は、MLRによって誘発される運動を抑制する。
歩行開始と転倒の失敗に苦しむPD患者の治療には、脚橋被蓋核(CNF)とPPNの背側部分を特に標的にする必要があるかもしれないことが示唆されている。
PDにおける皮質-小脳-脳幹経路の再編成
機能的ニューロイメージングアプローチを使用し、ヒトのSMAとPPN / MLR領域の間の強い機能的結合性を明らかにし、PD患者の凍結重症度と正の相関があった。
この強い接続性は、代償が原因であると考えられている。
逆に、STNとSMAの関係性は失われ、大脳基底核による歩行の自動制御の低下を反映している可能性があることが示唆される。
拡散テンソル画像を使用した研究により、PD患者の小脳とPPNの関係が明らかになった。
しかし、PD患者の冷凍庫は、小脳接続の欠如と前橋の皮質橋線維の可視性の増加を示した。
これらの所見は、歩行の病態生理学における皮質橋-小脳経路の重要性を強調している。
PDの冷凍庫では、歩行制御の自動実行に寄与する可能性のある小脳接続が損傷していることが考えられる。
読んでいただきありがとうございました。