
運動野は筋収縮を表現するのか、運動を表現するのか?
大脳皮質運動野はどのようにして行動を指令するのか?
解剖学的には哺乳類の大脳皮質は5b層の皮質脊髄路を通して脊髄に神経活動を伝え、脊髄が筋肉の動きを規定することで行動を起こすことが知られている。一方で、大脳皮質が行動に対してどのような影響を持つのかについては、2つの相異なる仮説があり、長らく論争が続いていた。
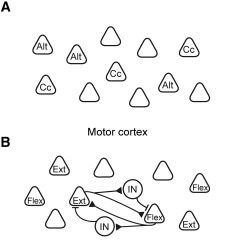
1つ目の仮説は、運動皮質が筋肉の共収縮や一方の筋肉の収縮と他方の筋肉の弛緩といった運動機能単位に対応した活動を示す神経集団を持つという仮説である(図1A)。古くは1874年に、サル脳の運動野を電気刺激することで、口を開けながら手を口元に運ぶといった動作が誘発されることが観察されている。この仮説が正しいとすると、例えば筋肉が共収縮する必要のある運動する物体をとどめておくタスク(Cc)や、対になった筋肉が相反する動きをする必要がある輪を手で回転させるタスク(Alt)を設計した際に、相異なる神経集団が活性化されることが予想される。
2つ目の仮説は、運動皮質が筋肉の弛緩や収縮に直接対応した活動を示す神経集団を持つという仮説である(図1B)。こちらも古く1870年には、イヌの大脳皮質の電気刺激により対則の筋肉が収縮することが報告されている。この仮説が正しいとすると、筋肉が共収縮する必要のあるタスクではそれぞれの筋肉の収縮に対応する神経集団が、筋肉が相反する動きをする必要があるタスクでは一方の筋肉の収縮(Flex)に対応する神経集団と他方の筋肉の弛緩(Ext)に対応する神経集団が活性化されることが予想される。
今回紹介する以下の論文では、上記のタスクとシリコンプローブ、2光子顕微鏡を用いた神経活動記録をマウスの大脳皮質運動野5b層の神経および皮質脊髄路の神経から行うことにより、これらどちらの仮説も正しくなく(個々のニューロンを筋肉の収縮や運動単位と関連付けることはできず)、運動皮質はその神経活動の共分散によって機能単位(タスク)に対応した情報を伝えているというモデルを提示した(図1C)。このモデルでは、運動皮質の神経集団のある次元における共分散が大きいと筋肉の共収縮を引き起こし、別の次元における共分散が大きいと筋肉の相反する動きを引き起こす。これは、運動野のタスク特異的な集団活動が様々な行動を制御しているという見方と合致する。

Warriner, Claire L., et al. "Motor cortical influence relies on task-specific activity covariation." Cell reports 40.13 (2022).
DOI: https://doi.org/10.1016/j.celrep.2022.111427

Methods
図1では本論文で開発された、筋肉の共収縮や相反する活動を必要とする行動パラダイムが説明されている。輪を回すタスクでは対になった筋肉が交互に活性化し、周期的に腕が動かされる(図1D-F)。一方で、外力に反して取手を特定の場所にとどめておくタスクでは筋肉が共収縮する(図1H-J)。活性化した筋肉が明るい赤で示され、特定のトライアルでの筋電図(EMG)と腕の位置の例(図1E,I)と全トライアルを平均した筋電図が示されている(図1F,J)。

Results
図3は、タスク依存的に強く活動する神経集団がないことを示すことで仮説1を否定している。マウス運動野の前腕運動に対応する領域(CFA)からシリコンプローブを用いてタスク中の神経活動を記録した。交互に筋活動が変化するタスクと筋肉が共収縮するタスクでの神経の平均発火率(図3B)、その対角線からの距離(図3D)、特異性指標(図3F)をプロットすると、多くの細胞はどちらのタスクにも特異的に活動せず、連続的に分布が変化することから、運動皮質の神経は特定のタスクに特化したサブ神経集団を持っているのではないと結論付けた。

図4は、タスクをまたいで筋活動と対応した活動を示すように神経集団をフィットすることができないことから、仮説2を否定している。過学習を防ぐためのL2正則化項が追加された線形回帰であるリッジ回帰を用いて、すべての神経活動から上腕三頭筋(Tri)と上腕二頭筋(Bi)の筋電図の活動を予測するモデルを作成すると、交互に筋活動が変化するタスク(Alt)でモデルを学習した場合は共収縮するタスク(Cc)の筋活動を高い精度で予測することができず(図4A,B)、逆に学習した場合も同様であった(図4C,D)。

図5は、神経の集団的活動はタスク特異的な共分散を持つという新たなモデルを提示している。神経の集団活動を主成分分析(PCA)を用いて次元削減すると、より多くの分散を説明できる順にPC1,PC2と次元が決定されていき、潜在空間でのダイナミクスはその集団活動に特異的な共分散の変化を示している。運動皮質の神経活動の共分散の意義を明らかにするために、それぞれのタスク時の神経活動を用いてPCA解析を行った(図5A)。神経集団の交互に筋活動が変化するタスクの1エポック目のデータを用いてPCAを行い、他のタスクでどれほどの分散を説明できるかを定量化すると、交互に筋活動が変化するタスクの2エポック目(同じタスク)の説明できる分散は大きかったが、他のタスクでは小さかった(図5B)。潜在空間での神経活動ダイナミクスを可視化すると、タスクごとに神経活動が分離されていることが分かった(図5F)。


Discussion
以上の結果より、著者は運動野の個々の神経の活動は直接タスクの情報と相関しているわけではなく、筋肉の活動と相関しているわけでもない(タスク細胞や筋活動細胞は存在しない)が、運動野の神経集団の活動の共分散がタスクの情報と相関していると結論付けた。
運動野の神経集団がタスク特異的な情報を共分散により伝えているとすると、今度はどのような上流の活動がこの活動を形成しているのか、下流にはどのように活動が伝えられているのかが疑問となる。下流については、皮質脊髄路の神経も運動野5b層と同様の活動を示すことが図7で示されており、タスク特異的な共分散の情報が脊髄の介在神経でデコードされ、運動神経に筋肉の弛緩や収縮に対応した情報を伝えていると考えられる。上流については、感覚野からの感覚フィードバックがタスクの情報を伝えることで運動野の活動を規定しているのではないかとディスカッションされている。
このような神経集団の活動の共分散によって情報をエンコードするというコーディング方法がなぜ進化したのか、単に各細胞が特定の情報をエンコードする場合に比べてどのような利点があるのかが気になった。
参考文献
新研究によれば、大脳皮質運動野は既存の2つの仮説を否定し、神経活動の共分散を用いて行動を指示する新モデルを提示。個々の神経が筋肉活動やタスクに直接関連するわけではなく、神経集団の共分散が行動を制御すると指摘。これにより、異なる行動タスクで神経活動の共分散が異なることが示された。
サムネイル画像の出典:https://doi.org/10.1016/j.celrep.2022.111427